Caracteres de las flores de la vid
Continuamos con la serie de post especializados en vid.
Estos post son destinados a mejorar las calidades descriptivas de las variedades que se cultivan y para el mejor entendimiento de las descripciones realizadas, es por tanto un ejercicio de ingenieria y lenguaje.
Haré una descripción de las cepas, sarmientos, hojas, flores, racimos y uvas.
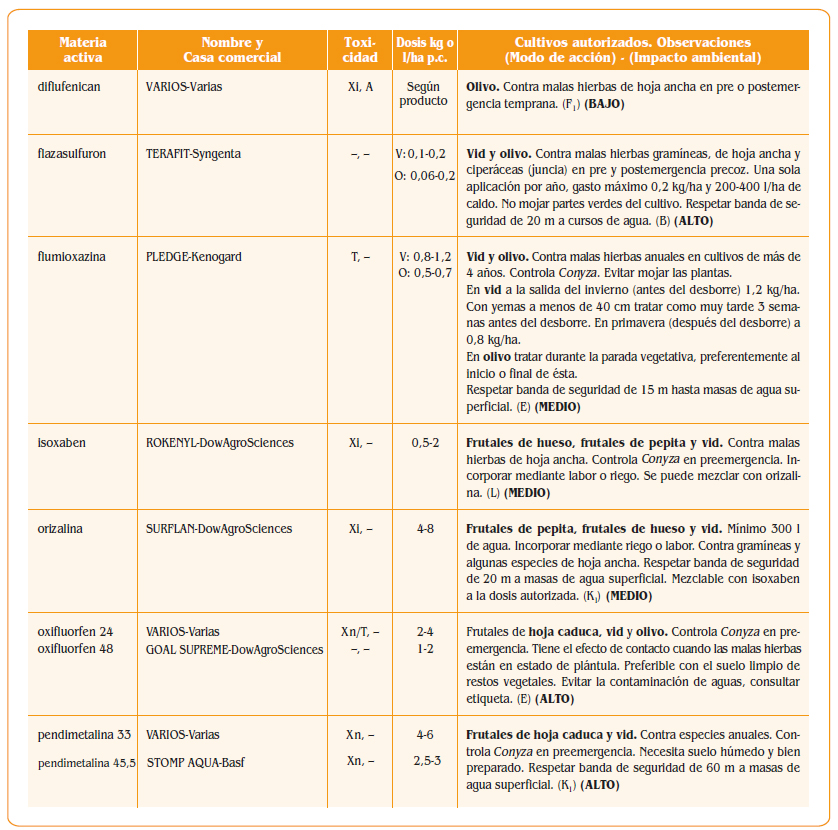
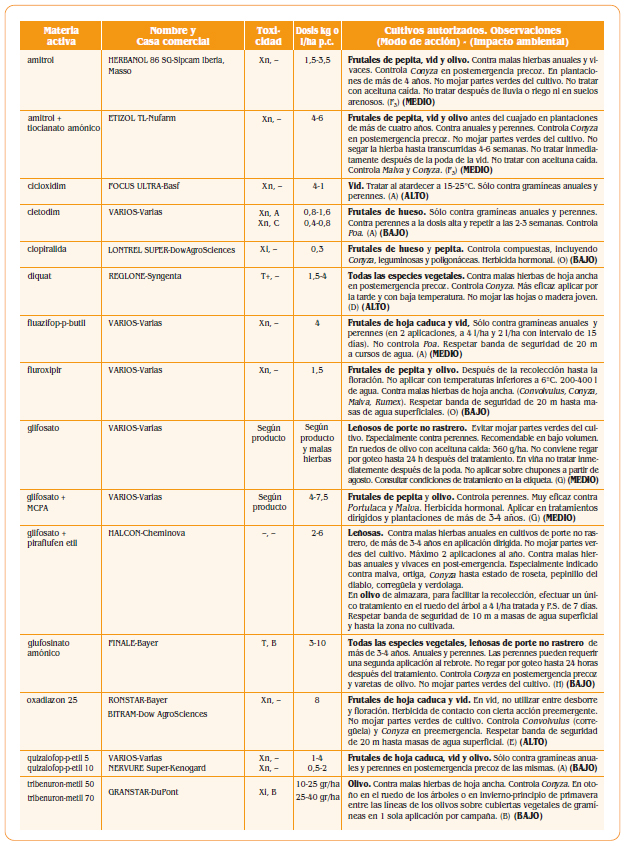
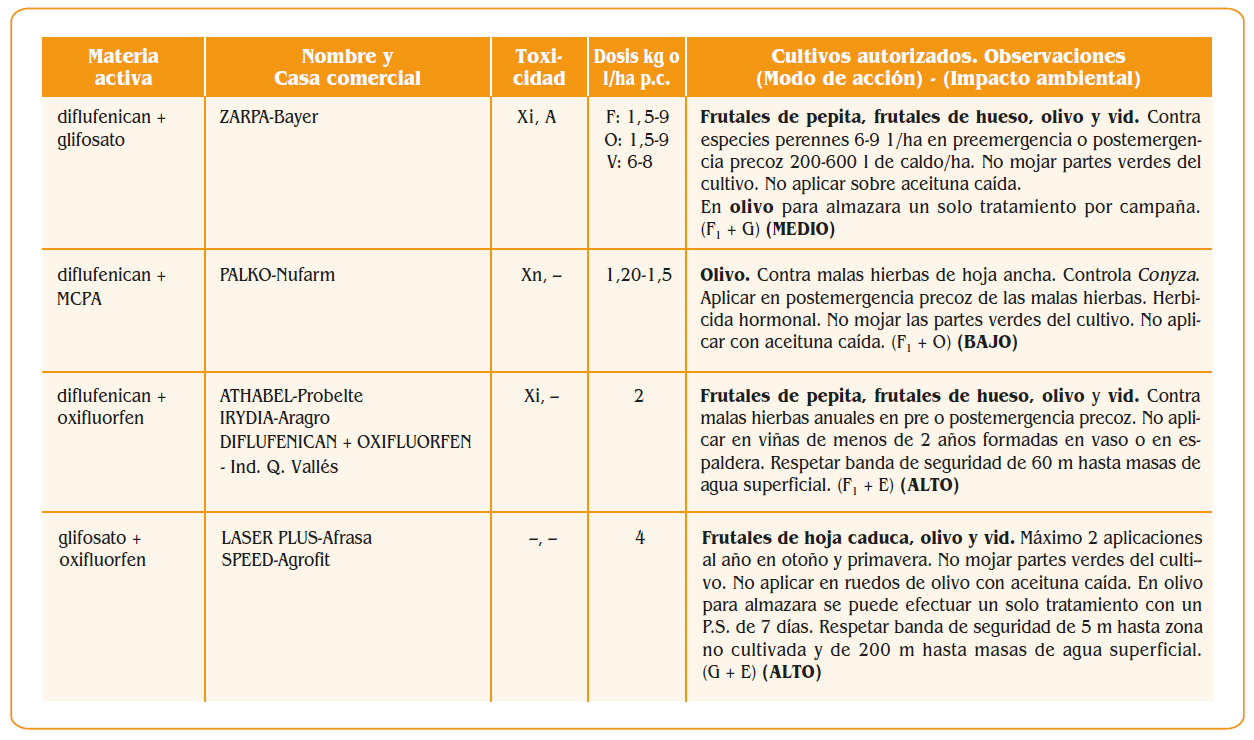
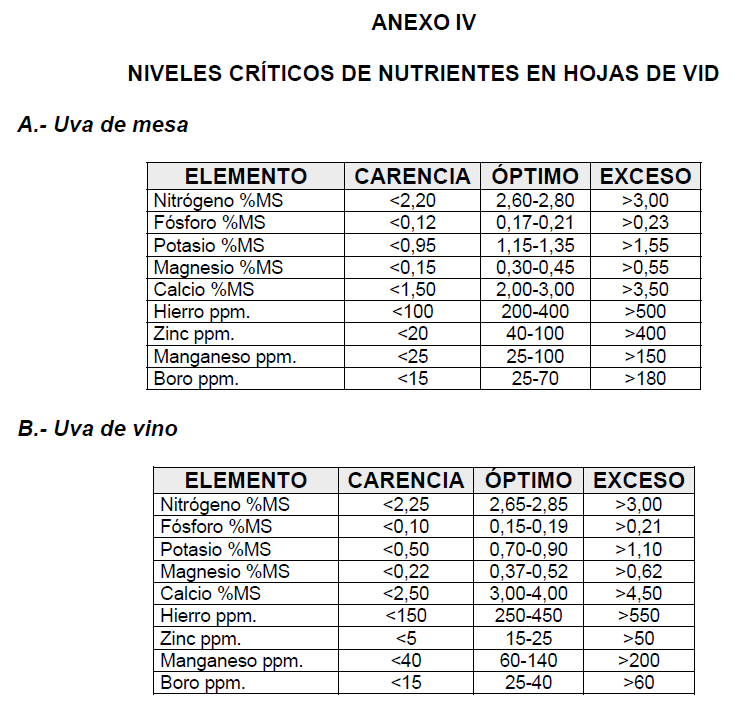
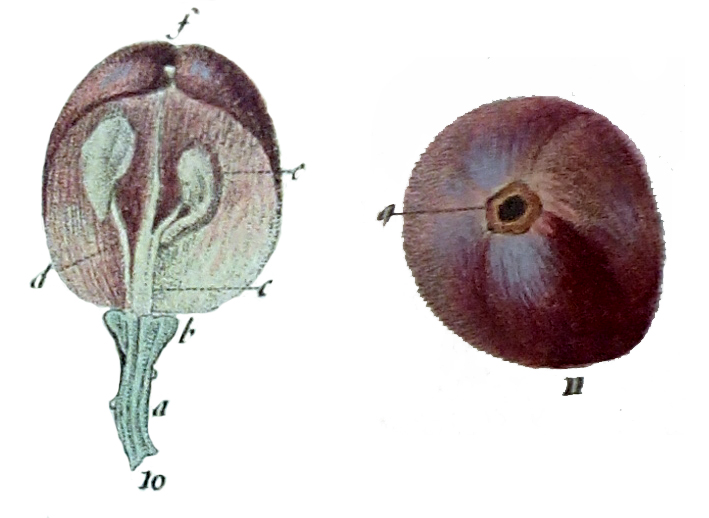
 7.a. – Peciolo
7.a. – Peciolo- 7.b. – Bractea que cubre el peciolo
- 7.c. – Caliz
- 8.a. – Peciolo
- 8.b. – Ovario
- 8.i. – Estigma
- 8.d. – Corola prroxima a caer
- 8.e. – Filamentos
- 8.f. – Anteras
- 8.g. – Dos glandulas de las cinco que rodean la base del ovario
- 9.a. – Peciolo verrugoso
- 9.c. – Caliz con algunas verrugas
- 9.d. – Disco bastante abultado
- 9.e. – Filamentos algo ondulados
- 9.f. – Anteras marchitas
- 9.j. – Parte del hollejo pegado
FLORES
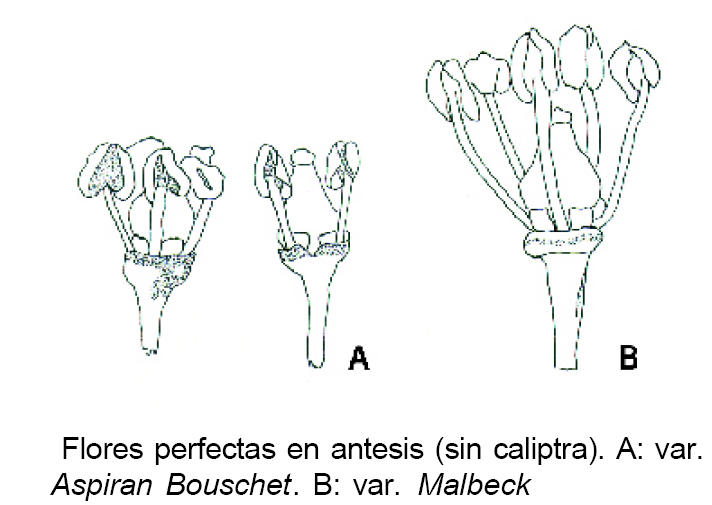
Las flores de la vid atendiendo a sus caracteristicas pueden ser:
- Con la Corola globosa antes de abrirse
- Con la Corola de figura de trompo
- Con todas las corolas caducas
- Con muchas corolas persistentes
- De cinco estambres
- De seis estambres
- De siete estambres
- De ocho estambres
- Con los estambres que caen antes de que pinte la uva
- Con los estambres que que persisten pintada ya la uva
- Con los estambres que que persisten madura ya la uva
- Con los estambres ensortijados después de marchitos
- Con los estambres ondeados después de marchitos
- Con los estambres estirados después de marchitos
- Con el disco muy abultado
- Con el disco bastante abultado
- Con el disco poco abultado
- Con el disco muy abultado
- Con el disco bastante abultado
- Con el disco poco abultado
- Caen muchas
- Caen bastantes
- Caen pocas
- Se marchitan muchas
- Se marchitan bastantes
- Se marchitan pocas
- Se desenvuelven aprisa
- Se desenvuelven lentamente
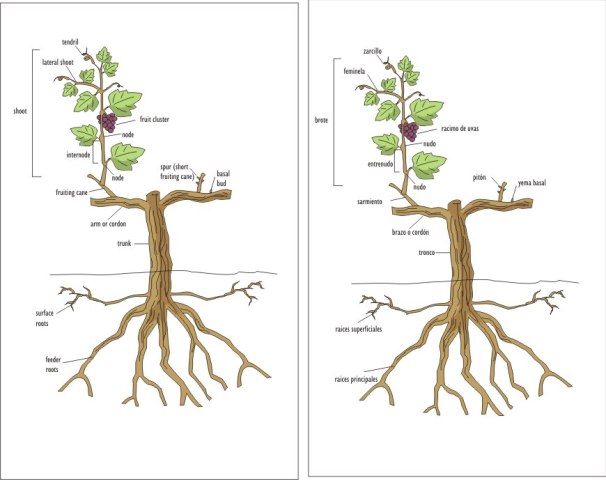
A continuacuión pongo este magnifico trabajo de Ed Hellman
Ed Hellman, Extensión Cooperativa de Texas
Yemas
Una yema es un punto de crecimiento que se desarrolla en la axila de la hoja, que es el área justo por encima del punto de conexión entre el peciolo y el brote. Esta yema que se desarrolla en esta área se describe en términos botánicos como una yema axilar. Es importante entender que hay una yema que se desarrolla en cada axila de la hoja de la vid, incluyendo las estípulas basales (hojas con forma de escamas). En la terminología vitícola, las dos yemas asociadas a una hoja se denominan yema normal y la yema pronta o anticipada. La yema normal es la verdadera yema axilar de la hoja, y la yema pronta o anticipada se forma en la axila de las estípulas de las yemas normales. Debido a su asociación de desarrollo, las dos yemas están situadas a cada lado de la axila de la hoja principal.

|
Una sección transversal de una yema dormida. Donde se pueden observar los tres conos vegetativos. |
Aunque la yema normal se ve como una estructura simple, en realidad es una yema compuesta, que contiene tres conos de crecimiento distintos. Estos se refieren comúnmente como conos primarios, secundarios y terciarios respectivamente dentro de una misma yema.La distinción entre las yemas secundarias y terciarias a veces es difícil de hacer cuando se observa la yema en forma visual y suele ser de poca importancia, por lo que es común referirse a los dos conos de crecimiento más pequeños como las yemas secundarias. Estos tres conos de crecimientos están juntos dentro de un grupo externo de escamas de protección dentro de la yema compuesta. A medida que la yema se desarrolla, se sigue el patrón de nomenclatura de las yemas en el brote: el cono de crecimiento primario es la yema axilar de las yemas laterales, los conos secundarios y terciarios de crecimiento son las yemas axilares de las dos primeras estípulas del cono primario de crecimiento.
Las yemas dormidas o latentes son el centro de atención durante la poda de invierno, ya que contienen los primordios de racimos florales (son el potencial de producción de fruta para la próxima temporada). Se llama latente para reflejar el hecho de que normalmente no crecen en la misma temporada en que se desarrollan.
Las yemas latentes se inician el año anterior a su crecimiento como un brote. Durante la temporada anterior, se someten a un desarrollo considerable. Los tres conos de crecimiento de la yema compuesta producirán brotes rudimentarios que finalmente contendrán los primordios (órganos en sus primeras etapas de desarrollo) de los mismos componentes básicos que conforman un brote adulto: hojas, zarcillos, y en algunos casos racimos florales. El cono primario se desarrolla primero, por lo que es el más grande y el más desarrollado en el momento en que la yema se aletarga. Si se produce bajo condiciones ambientales y de crecimiento favorables, contendrá primordios de racimo florales antes del final de la temporada de crecimiento. Los primordios de racimo florales representan por lo tanto el potencial de fructificación de la yema para la temporada siguiente. Como reflejo de la secuencia de desarrollo, los conos secundarios y terciarios son cada vez más pequeños y menos desarrollados. Por lo general serán menos fructíferos (tienen menos racimos y más pequeños) que el cono primario. La fecundidad de las yemas (potencial para producir fruta) es una función de la variedad, condiciones ambientales y prácticas de producción vitícola. Yemas latentes que se desarrollan en condiciones desfavorables (sombra en una canopia con denso follaje, mala alimentación, etc.) producen menos primordios de racimo florales para la temporada siguiente.
En la mayoría de los casos, sólo la yema primaria crece, produciendo el brote principal en la temporada siguiente. La yema secundaria puede ser considerada como un «sistema de copia de seguridad» de la vid, normalmente, sólo crece cuando el brote principal o brotes jóvenes han sufrido daños, a menudo por congelamiento o heladas en primavera. Sin embargo, bajo ciertas condiciones, tales como podas severas, la destrucción de una parte de la vid, o la deficiencia de boro, es posible que dos o los tres conos de crecimiento produzcan brotes en la primavera (Winkler et al., 1974). Las yemas terciarias proporcionan una copia de seguridad adicional en caso que las yemas primarias y secundarias estén dañadas, pero por lo general no tienen racimos florales y por lo tanto ningún fruto. Si sólo el tallo primario crece, las yemas secundarias y terciarias siguen vivas, pero latente en la base del brote.
La yema pronta crecerá en la actual temporada, pero el crecimiento puede cesar de pronto después de la formación de las estípulas o puede continuar, produciendo un brote lateral de longitud variable. Independientemente del grado de desarrollo de las yemas laterales, una yema compuesta se desarrolla en la estípula basal, formando la yema dormida. Los brotes laterales a veces producen racimos florales y frutas, que se conoce como «racimas». Sin embargo, debido a que estos se desarrollan más tarde en la temporada, «las racimas» no maduran totalmente en muchas zonas del país. Si una yema pronta no crece en la temporada actual morirá con los primeros fríos.
Flores y frutos
En un brote fructífero por lo general se producen de uno hasta tres racimos de flores (inflorescencias) dependiendo de la variedad. Los racimos florales se desarrollan en el lado opuesto al de las hojas, generalmente entre el tercer y el sexto nudo desde la base del brote, dependiendo de la variedad. Si se desarrollan tres racimos florales, dos de ellos se desarrollan en en nudos adyacentes, el siguiente racimo floral se desarrollará en un nudo superior dejando un nudo de por medio sin racimo. El número de racimos de florales en un racimo depende de la variedad de uva y de las condiciones de desarrollo de la temporada anterior en la cual la yema normal (que produjo el brote principal) se desarrolló. Un racimo puede contener varios cientos de flores individuales, dependiendo de la variedad.


Yemas y flores. Un yema compuesta con la yema primaria, secundaria y terciaria (L), y las flores desde la formación a la caída de la caliptra hasta la pre-fertilización.
La flor de la planta de uva no tiene pétalos visibles, en cambio, los pétalos se funden en una estructura verde denominada caliptra, pero comúnmente conocida como la capucha o tapa. La caliptra incluye los órganos reproductores y otros tejidos dentro de la flor. Una flor se compone de un solo pistilo (órgano femenino) y cinco estambres, cada uno con una antera en la punta (órgano masculino). El pistilo es aproximadamente de forma cónica, con la base mucho mayor que la parte superior, y el extremo (llamado el estigma) ligeramente acampanado. La base del pistilo es el ovario, y consta de dos compartimentos internos, cada una con dos óvulos que contiene un saco embrionario con un solo huevo. Las anteras producen muchos granos de polen amarillos, que contienen los espermatozoides. Vides silvestres, patrones o portainjertos (y unas pocas variedades cultivadas, tales como “St. Pepin”) poseen flores “pistiladas” (femeninas) o flores “estaminadas” (masculinas) por lo que la planta entera es masculina o femenina. Viñas con flores femeninas pistiladas necesitan de viñas cercanas con flores estaminadas o hermafroditas para la producción de fruta. La mayoría de las variedades de vid comerciales tienen flores hermafroditas, es decir, los dos componentes masculinos y femeninos.
Una flor individual se muestra con sus piezas florales etiquetadas.
El período de tiempo durante el cual las flores están abiertas (la caliptra ha caído) se llama floración y puede durar de 1 a 3 semanas dependiendo de las condiciones climáticas. Viticultores de diversas maneras se refieren a la plena floración como la etapa en que se han abierto o caído aproximadamente entre el 50% y dos tercios de las caliptras de las flores. La floración se produce normalmente entre 50 y 80 días después de la brotación.
Etapas de la Floración
Cuando las flores individuales en un racimo floral se abren se ven diferentes a las flores de la mayoría de flores de otras plantas. La caliptra se separa de la base de la flor, se sale, y por lo general se cae, dejando al descubierto el pistilo y las anteras. Las anteras liberan su polen ya sea antes o después de la caída de la caliptra. Los granos de polen caen al azar sobre el estigma del pistilo, lo que permite la polinización. Múltiples granos de polen pueden germinar y se desarrolla un tubo llamado tubo polínico desde el estigma hasta el óvulo, donde un espermatozoide se une con un óvulo para formar un embrión. El éxito de la unión se denomina fertilización, y el posterior crecimiento de las bayas se llama «cuaje». La baya se desarrolla a partir de los tejidos del pistilo, sobre todo el ovario. El óvulo, junto con su embrión se convierte en la semilla.
 |
Una inflorescencia con casi el 100% de las caliptras caídas. |
Debido a que hay cuatro óvulos por flor, hay un potencial máximo de cuatro semillas por baya. Condiciones ambientales desfavorables durante la floración, como el clima frío y lluvioso, puede reducir el cuajado (número de bayas) y semillas por baya, lo que afecta el tamaño de la baya. El tamaño de las bayas se relaciona con el número de semillas dentro de la baya, y muy pocas semillas lleva a pequeñas bayas. Sin embargo, el tamaño de las bayas también puede estar influenciada por las condiciones ambientales, prácticas de manejo y gestión del agua. Algunos frutos inmaduros pueden ser retenidos por el racimo sin completar su crecimiento y desarrollo normal, un fenómeno conocido como «corrimiento» (Mullins et al., 1992).
Referencias:
Mullins, M. G., A. Bouquet, E. y L. Williams. 1992. Biología de la Vid. Cambridge University Press.
Revisado por Tim Martinson, de la Universidad de Cornell y Patty Skinkis, de la Universidad Estatal de Oregon Traducido por Juan Pablo Valot
A continuación pondré este gran trabajo que complementa el articulo completamente.
FLOR Y FRUTO DE VID (Vitis vinifera L.) Micrografía aplicada a Viticultura y Enología
Claudia Victoria Lúquez * Juan Carlos Formento **
Estructura de la flor
El pedicelo de la flor presenta epidermis cutinizada monoestratificada (un estrato de células aplanadas o tabulares de pared tangencial externa engrosada), colénquima angular (banda debajo de la epidermis, con células de pared primaria irregularmente engrosada) y parénquima cortical (células grandes de pared primaria delgada, con cloroplastos y almidón). Presenta cinco haces colaterales vasculares en su base, que luego se separan para formar nuevos hacecillos dirigiéndose a las piezas florales.
 Se observan idioblastos (células que se diferencian marcadamente de las restantes de un tejido) llamados células rafidíferas. Son fácilmente visibles por sus enormes vacuolas ocupadas por los ráfides (manojos de rafidios o cristales de oxalato de calcio con forma de aguja) (foto 1).
Se observan idioblastos (células que se diferencian marcadamente de las restantes de un tejido) llamados células rafidíferas. Son fácilmente visibles por sus enormes vacuolas ocupadas por los ráfides (manojos de rafidios o cristales de oxalato de calcio con forma de aguja) (foto 1).
Los sépalos poseen dos epidermis monoestratificadas, varias capas de parénquima fundamental y el sistema vascular representado por los haces vasculares. No muestran diferenciación entre parénquimas esponjoso y en empalizada. Su color verde se debe a que sus células poseen cloroplastos.
 La caliptra de la vid es la corola dialipétala, con los pétalos libres al principio y soldándose al finalizar su desarrollo para formar una capucha que cubre a las anteras y al gineceo (foto 2).
La caliptra de la vid es la corola dialipétala, con los pétalos libres al principio y soldándose al finalizar su desarrollo para formar una capucha que cubre a las anteras y al gineceo (foto 2).
La base de cada pétalo presenta una capa de pequeñas células poco diferenciadas en el lugar donde luego se producirá la absición. La caliptra se separa desde su base y cae en conjunto en el momento de la antesis. Existen flores denominadas en estrella con los pétalos separados en el ápice y la corola que no forma caliptra. Los pétalos presentan epidermis abaxial y adaxial monoestratificadas (con estomas y cutícula listada), parénquima fundamental constituido por 3-5 capas de células y un haz vascular.
El estambre contiene los sacos polínicos en cuyo interior se encuentran los granos de polen. Los estambres son opuestos a los pétalos (opositipétalos) y libres entre sí. Constan de dos partes bien delimitadas: antera (parte superior fértil) y filamento (parte basal estéril).
La antera es dorsifija y tetraloculada (con cuatro cavidades). Tiene dos tecas, con dos sacos polínicos cada una.
 El conectivo es la parte media estéril de la antera y en la vid adopta característicamente la forma de una bifurcación que conecta ambas tecas, provocando que la antera sea bilobada (foto 3).
El conectivo es la parte media estéril de la antera y en la vid adopta característicamente la forma de una bifurcación que conecta ambas tecas, provocando que la antera sea bilobada (foto 3).
El filamento presenta una epidermis similar a la de la corola, parénquima fundamental y un haz vascular anficribal (el floema rodea al xilema), circundado por el parénquima fundamental.
 El saco polínico está constituido por diferentes capas de tejidos que forman su pared y por el tejido esporógeno (dentro del lóculo). Las capas parietales son, de afuera hacia dentro: (foto 4).
El saco polínico está constituido por diferentes capas de tejidos que forman su pared y por el tejido esporógeno (dentro del lóculo). Las capas parietales son, de afuera hacia dentro: (foto 4).
- • Epidermis formada por células tabulares que se comprimen a la madurez de la antera.
- • Endotecio o capa fibrosa: la capa parietal típica de la antera relacionada con el mecanismo de dehiscencia. Sus células tienen la pared tangencial interna engrosada en forma homogénea y las paredes radiales con engrosamientos de pared secundaria en forma de barras, bandas o filetes lignificados, alternadas con franjas que poseen solamente pared primaria. En la región del estomio (pequeña ranura longitudinal) el endotecio no desarrolla espesamientos secundarios. En la dehiscencia de la antera, cuando el endotecio se contrae tangencialmente, el estomio se rompe permitiendo la salida del polen.
- • Capa media, células parenquimáticas que aparecen muy aplastadas en la antera madura.
- • Tapete secretor o glandular: capa de tejido más interna, especializada en la nutrición de las microesporas y formación de la exina del grano de polen.
 El gineceo forma un solo cuerpo o pistilo. Posee una parte inferior fértil (ovario) y una parte superior estéril (estigma y estilo). Es bicarpelar (con dos carpelos) y gamocarpelar (con los carpelos unidos). Cada carpelo se pliega por su nervadura media, de manera que la cara superior mira hacia el interior y se suelda a sí mismo (margen contra margen, determinado un lóculo en su interior). Los márgenes carpelares (donde están las placentas) se ubican en el centro del ovario (en el axis o eje). Por lo tanto, la placentación en vid es marginal axial. A su vez, ambas hojas carpelares se unen entre sí por su cara inferior formando un septum (tabique) que divide al ovario en dos cavidades. El gineceo es, por lo tanto, bilocular (foto 5).
El gineceo forma un solo cuerpo o pistilo. Posee una parte inferior fértil (ovario) y una parte superior estéril (estigma y estilo). Es bicarpelar (con dos carpelos) y gamocarpelar (con los carpelos unidos). Cada carpelo se pliega por su nervadura media, de manera que la cara superior mira hacia el interior y se suelda a sí mismo (margen contra margen, determinado un lóculo en su interior). Los márgenes carpelares (donde están las placentas) se ubican en el centro del ovario (en el axis o eje). Por lo tanto, la placentación en vid es marginal axial. A su vez, ambas hojas carpelares se unen entre sí por su cara inferior formando un septum (tabique) que divide al ovario en dos cavidades. El gineceo es, por lo tanto, bilocular (foto 5).
 El estigma -superficie que recibe al grano de polen- es del tipo húmedo o glandular, con una epidermis glandular papilada, constituida por células alargadas unidas por sus extremos formando papilas (cordones filamentosos) de aproximadamente 20 células de longitud. Las células epidérmicas y subepidérmicas secretan una solución azucarada constituida por compuestos lipídicos y fenólicos, que tapiza en forma de película el estigma y el canal estilar luego de la antesis (foto 6).
El estigma -superficie que recibe al grano de polen- es del tipo húmedo o glandular, con una epidermis glandular papilada, constituida por células alargadas unidas por sus extremos formando papilas (cordones filamentosos) de aproximadamente 20 células de longitud. Las células epidérmicas y subepidérmicas secretan una solución azucarada constituida por compuestos lipídicos y fenólicos, que tapiza en forma de película el estigma y el canal estilar luego de la antesis (foto 6).
 El estilo es corto. Posee un canal estilar (conducto por el cual se desplaza el tubo polínico) que va desde el estigma hasta cada uno de los lóculos. El estilo consiste de epidermis monoestratificada, parénquima fundamental con espacios intercelulares y tejido estigmatoide (revestimiento interno glandular del canal estilar, que continúa la epidermis glandular del estigma). Estos tejidos tienen idioblastos con ráfides (foto 7).
El estilo es corto. Posee un canal estilar (conducto por el cual se desplaza el tubo polínico) que va desde el estigma hasta cada uno de los lóculos. El estilo consiste de epidermis monoestratificada, parénquima fundamental con espacios intercelulares y tejido estigmatoide (revestimiento interno glandular del canal estilar, que continúa la epidermis glandular del estigma). Estos tejidos tienen idioblastos con ráfides (foto 7).
 El tejido estigmatoide se prolonga en el lóculo recubriendo ambos lados de la placenta y el funículo del óvulo. El tubo polínico crece intercelularmente en el estigma y en el canal estilar emerge dentro del lóculo, avanza por la superficie de la placenta y del funículo, y entra por la micrópila del óvulo. La pared del ovario presenta tres capas celulares (foto 8):
El tejido estigmatoide se prolonga en el lóculo recubriendo ambos lados de la placenta y el funículo del óvulo. El tubo polínico crece intercelularmente en el estigma y en el canal estilar emerge dentro del lóculo, avanza por la superficie de la placenta y del funículo, y entra por la micrópila del óvulo. La pared del ovario presenta tres capas celulares (foto 8):
- • Epidermis abaxial o externa, monoestratificada: sus células poseen gotitas refractivas de polifenoles antes de la antesis. Hay estomas en la proporción de 1-2 /mm2.
- • Mesófilo con un estrato de 9-11 filas de espesor de células parenquimáticas, a ambos lados de la red de haces vasculares. Hasta el momento de la antesis, el ovario está cubierto por la caliptra de pétalos, que impide la llegada de la luz solar directa. Esto explicaría el hecho de que el contenido de clorofila presente en el mesófilo es muy bajo cuando cae la caliptra, aumenta drásticamente cuando se expone el ovario a la luz solar y va disminuyendo paulatinamente durante el proceso de maduración de la baya, con la consiguiente pérdida de color verde. Este fenómeno coincide con el incremento del contenido de los monoterpenos responsables del aroma en la baya. Los haces vasculares penetran el carpelo provenientes del receptáculo: un haz dorsal (nervadura media) y dos haces ventrales que se desplazan por los márgenes de la hoja carpelar y alimentan a las placentas ubicadas en la zona axial. Otras ramificaciones de los haces vasculares dorsales y ventrales forman una red periférica de haces vasculares. Todos convergen finalmente en el estilo.
- • Epidermis adaxial o interna, con 1-2 capas de células planenquimáticas, cuyas vacuolas están aparentemente libres de polifenoles. El estrato más interno contiene drusas (cristal de oxalato de calcio, compuesto de cristales menores que se proyectan desde su superficie).
 El disco de nectarios, formado por cinco glándulas, se encuentra en la base del ovario de las flores hermafroditas, funcionalmente femeninas y masculinas. Puede ser funcional o no, pero produciría el olor típico de la inflorescencia de la vid. Los nectarios derivarían filogenéticamente de la transformación de un verticilo de estambres. Experiencias realizadas sobre la percepción olfativa en seres humanos han demostrado que el olor característico de flores de vid es más fuerte en las flores masculinas que en las femeninas. En algunas flores se forma un segundo disco poco antes de la antesis, entre el cáliz y la corola, y continúa creciendo luego de la fertilización (foto 9).
El disco de nectarios, formado por cinco glándulas, se encuentra en la base del ovario de las flores hermafroditas, funcionalmente femeninas y masculinas. Puede ser funcional o no, pero produciría el olor típico de la inflorescencia de la vid. Los nectarios derivarían filogenéticamente de la transformación de un verticilo de estambres. Experiencias realizadas sobre la percepción olfativa en seres humanos han demostrado que el olor característico de flores de vid es más fuerte en las flores masculinas que en las femeninas. En algunas flores se forma un segundo disco poco antes de la antesis, entre el cáliz y la corola, y continúa creciendo luego de la fertilización (foto 9).
Grano de polen El grano de polen normal es tricolporado: con tres surcos, más anchos en el ecuador que en los polos. Cada surco posee un poro redondeado en su parte media. Tiene simetría radial y es isopolar, elíptico alargado en vista ecuatorial, y triangular en vista polar, con los ángulos surcados y los lados cóncavos. Al absorber agua los ángulos se dilatan y su forma se hace hexagonal por dilatación. La pared del grano de polen maduro consiste de una gruesa capa externa (exina) y una delgada capa interna (intina). En el momento de la dehiscencia de la antera, el grano de polen es binucleado y la exina ha desarrollado los tres surcos longitudinales y los tres poros germinativos. Los poros -a través de los cuales emerge el tubo polínico son áreas delgadas de la exina donde se visualiza la intina, compuesta de pectina y celulosa. La pared está constituida por esporopolenina (polímeros oxidativos de carotenoides y ésteres carotenoides) notablemente resistente a las altas temperaturas y a los agentes químicos. También presenta enzimas y tipos alergénicos, responsables de las reacciones de incompatibilidad durante la polinización. El tipo de polinización en la vid es discutido y sería de tres tipos: anemófila, entomófila y autopolinización. Un alto porcentaje de vides serían autógamas (pudiendo darse la autopolinización en la yema), el resto entomófilas alógamas y sólo unas pocas anemófilas.
Óvulo  El óvulo de Vitis es anátropo, con nucela sólida y dos tegumentos (foto 10).
El óvulo de Vitis es anátropo, con nucela sólida y dos tegumentos (foto 10).
La nucela es el cuerpo central del óvulo, con células vegetativas que rodean el saco embrionario. Éste es del tipo normal o tipo Polygonum (consta de siete células: célula huevo, dos sinérgidas, una célula central con dos núcleos y tres antípodas).
El funículo del óvulo es el pie que lo une a la placenta. Un haz vascular lo recorre hasta la chalaza. Tiene xilema con traqueidas con engrosamientos secundarios espiralados lignificados, y floema con células cribosas. Tiene una vaina de esclerénquima cerca de la chalaza.
El tegumento externo consiste de 2-9 capas de células, la más externa con células taníferas.
El tegumento interno posee 2-3 capas de células. La capa más externa posee cutícula, mientras que las células de la más interna se encuentran llenas de tanino.
En la micrópila, el tegumento interno se proyecta por fuera del tegumento externo, formando el endostomio (collar alrededor de la micrópila).
Fruto Es el ovario desarrollado luego de la fecundación. Se trata de una baya, un fruto carnoso pluriseminado, indehiscente a la madurez. También son carnosos los tabiques y las placentas.
El pericarpo, o pared del fruto, en la vid está dividida en tres capas: epicarpo (epidermis externa e hipodermis externa), mesocarpo (pared externa y pared interna) y endocarpo (hipodermis y epidermis internas).
La cera epicuticular compone la característica pruina sobre la epidermis cuticularizada de los frutos Está formada por delgadas fibrillas que sirven de alojamiento a las levaduras responsables de la fermentación del mosto, así como también diversos gérmenes de enfermedades criptogámicas. Se inicia en floración como pequeñas plaquetas verticales, sobre y entre los cordones cuticulares. A la madurez tienen 0,1 μ de diámetro, con bordes lobulados, desordenadamente superpuestas formando una capa hidrofóbica de varios micrones de espesor. Estas plaquetas son de naturaleza marcadamente hidrofóbica y totalizan 0,09-0,11 mg/cm2 (las hojas de la misma variedad contienen 1/10 de ese valor). Entre ellas se forma una compleja red de canales capilares, de longitud mucho mayor que el espesor de la capa de cera, que reduce notablemente la transpiración cuticular. Para acelerar la deshidratación de las pasas de uva, se las sumerge en soluciones comerciales que llenan los largos y angostos canales capilares hidrofóbicos, desalojando el aire entre dichas plaquetas y facilitando la transferencia de vapor de agua a través de la capa de cera.
Los fungicidas y detergentes afectarían la inclinación y la forma de las plaquetas de cera. La cera epicuticular está constituida por ceras blandas (30 %) en la zona externa, formando las plaquetas y ceras duras (70 %) más abundantes en las capas internas próximas a la cutícula. Las ceras blandas son una mezcla de ésteres de ácidos simples de larga cadena combinados con monoalcoholes alifáticos, y pequeñas cantidades de aldehidos, ésteres, ácidos grasos, hidrocarburos y ácido oleanólico. Las ceras duras están constituidas principalmente por ácido oleanólico (60 %) y lípidos (10 %) (24).
La cutícula es la zona que primero entra en contacto con los patógenos y juega un papel importante en la defensa de la planta. Es una capa acelular traslúcida de 1,6-3,8 μ de espesor, con cordones superficiales paralelos al eje del fruto. Esta constituida en un 65 % por cutina, un poliéster lipídico insoluble, elástico y resistente a la degradación térmica. Comprende una capa externa (la lamela cuticular, región libre de celulosa, con lamelas separadas por cutina y cera), una capa interpuesta (la región reticulada comprendida por cutina y cera atravesadas por fibrillas de celulosa) y una capa interna (la lamela péctica).
La piel, epicarpo u hollejo (película) de la uva está constituida por la cutícula que lo cubre, la epidermis externa, la hipodermis subyacente y algunas capas de células parenquimáticas subyacentes. Esta película externa de la baya presenta 2 tipos de células en los preparados microscópicos: células con precipitados granulosos o «células taníferas» y células desprovistas de precipitados coloreados. La epidermis externa consiste de células planenquimáticas con la cara tangencial externa más gruesa y vacuolas con algunos precipitados fenólicos, antocianos y taninos.  Cuando la baya madura, el número de estomas/mm2 disminuye y quedan aislados por lenticelas corchosas, lo que contribuye a una disminución del intercambio gaseoso, así como de la transpiración y la fotosínteis en la baya madura. La hipodermis externa tiene un espesor de 100-250 μ y está formada por 1-17 hileras de células colenquimatosas (de paredes primarias gruesas e irregulares), aplanadas y alargadas en sentido longitudinal. Contienen polifenoles vacuolares, flavonoides en las uvas blancas y antocianinas en las uvas tintas, que se ven bajo cuatro formas en los preparados: gotitas refractivas, masas densas amorfas, precipitados granulares finos a gruesos y masas periféricas (foto 11).
Cuando la baya madura, el número de estomas/mm2 disminuye y quedan aislados por lenticelas corchosas, lo que contribuye a una disminución del intercambio gaseoso, así como de la transpiración y la fotosínteis en la baya madura. La hipodermis externa tiene un espesor de 100-250 μ y está formada por 1-17 hileras de células colenquimatosas (de paredes primarias gruesas e irregulares), aplanadas y alargadas en sentido longitudinal. Contienen polifenoles vacuolares, flavonoides en las uvas blancas y antocianinas en las uvas tintas, que se ven bajo cuatro formas en los preparados: gotitas refractivas, masas densas amorfas, precipitados granulares finos a gruesos y masas periféricas (foto 11).
Hay idioblastos con rafidios de tartrato de Ca, oxalato de Ca o tartrato de potasio (15). Existen cultivares en las cuales la piel de la baya se separa fácilmente de la pulpa debido a que las células parenquimáticas ubicadas bajo la hipodermis poseen paredes mucho más delgadas y han perdido parte de las sustancias pécticas que la unen a otras.
 El mesocarpo o pulpa se divide topográficamente en mesocarpo externo a los haces vasculares periféricos, el grosor del cual varía con la cultivar y mesocarpo interno a los haces vasculares. Presenta 25-30 capas de células parenquimáticas, con paredes primarias celulósicas delgadas, vacuola central, isodiamétricas, más alargadas a medida que se acercan a la hipodermis, y con abundantes espacios intercelulares (foto 12). En el momento de la antesis, son muy ricas en polifenoles, aunque algunas células más pequeñas del mesocarpo nunca muestran polifenoles en sus vacuolas. Dos semanas después pierden los polifenoles, por lo que la pulpa normalmente no tiene color. Cuando no se produce esta pérdida de color, manteniéndose la pulpa coloreada en el momento de la madurez, las cepas se denominan tintoreras. Cuando la baya está verde, tiene cloroplastos, taninos y numerosos idioblastos con ráfides de oxalato de calcio. El incremento de volumen está acompañado por una elevación en la tasa de acumulación de glucosa y fructosa. El mesocarpo representa el 64 % del volumen final de la baya.
El mesocarpo o pulpa se divide topográficamente en mesocarpo externo a los haces vasculares periféricos, el grosor del cual varía con la cultivar y mesocarpo interno a los haces vasculares. Presenta 25-30 capas de células parenquimáticas, con paredes primarias celulósicas delgadas, vacuola central, isodiamétricas, más alargadas a medida que se acercan a la hipodermis, y con abundantes espacios intercelulares (foto 12). En el momento de la antesis, son muy ricas en polifenoles, aunque algunas células más pequeñas del mesocarpo nunca muestran polifenoles en sus vacuolas. Dos semanas después pierden los polifenoles, por lo que la pulpa normalmente no tiene color. Cuando no se produce esta pérdida de color, manteniéndose la pulpa coloreada en el momento de la madurez, las cepas se denominan tintoreras. Cuando la baya está verde, tiene cloroplastos, taninos y numerosos idioblastos con ráfides de oxalato de calcio. El incremento de volumen está acompañado por una elevación en la tasa de acumulación de glucosa y fructosa. El mesocarpo representa el 64 % del volumen final de la baya.
 El endocarpo tiene 2-3 capas de células y posee una epidermis interna (junto al lóculo) con células más pequeñas que las del mesocarpo, paredes gruesas y vacuolas sin polifenoles. Hacia el endocarpo hay una hipodermis interna, un estrato de células que contienen drusas y están aparentemente libres de polifenoles (foto 13).
El endocarpo tiene 2-3 capas de células y posee una epidermis interna (junto al lóculo) con células más pequeñas que las del mesocarpo, paredes gruesas y vacuolas sin polifenoles. Hacia el endocarpo hay una hipodermis interna, un estrato de células que contienen drusas y están aparentemente libres de polifenoles (foto 13).
El septum (tabique) crece rodeando las semillas o llenando los lóculos donde las semillas han abortado. Sus células son irregulares en tamaño y forma.
El sistema vascular presenta un haz vascular dorsal (nervadura media) con bandas vasculares periféricas (red periférica) y haces vasculares ventrales (en los márgenes de la hoja carpelar). Las haces vasculares tienen xilema con traqueidas y vasos con engrosamientos secundarios espiralados, floema con células cribosas, y parénquima de células elongadas. Los extremos rotos de las bandas vasculares constituyen el pincel cuando el pedúnculo es arrancado de la baya madura.
 El pedúnculo posee (foto 14): epidermis con unos pocos estomas; varias capas de células colenquimáticas (subepidérmicas); células parenquimáticas, de mayor volumen, con cloroplastos y almidón; idioblastos con ráfides; haces vasculares colaterales, cambium, y médula. El grosor del pedúnculo, así como el tamaño de la baya madura se incrementa con el número de semillas. La zona de abscisión se origina en la base del pedúnculo (si no hay cuajado de fruto) y en la base la baya si ésta llega a la madurez.
El pedúnculo posee (foto 14): epidermis con unos pocos estomas; varias capas de células colenquimáticas (subepidérmicas); células parenquimáticas, de mayor volumen, con cloroplastos y almidón; idioblastos con ráfides; haces vasculares colaterales, cambium, y médula. El grosor del pedúnculo, así como el tamaño de la baya madura se incrementa con el número de semillas. La zona de abscisión se origina en la base del pedúnculo (si no hay cuajado de fruto) y en la base la baya si ésta llega a la madurez.
Los granos de uva que no poseen semillas se llaman apirénicos. En la apirenia corintiana las uvas son partenocárpicas. Sus óvulos han abortado sin saco embrionario y con un solo tegumento o con saco embrionario defectuoso.
 En la apirenia sultanina las uvas son estenospérmicas con pepitas mal desarrolladas, donde hubo fecundación pero el embrión y el endosperma cesan su evolución. En algunas flores normales, el ovario puede tener 1-2 lóculos más de lo normal, que también contienen óvulos, pero solamente uno o dos lóculos son funcionales (foto 15).
En la apirenia sultanina las uvas son estenospérmicas con pepitas mal desarrolladas, donde hubo fecundación pero el embrión y el endosperma cesan su evolución. En algunas flores normales, el ovario puede tener 1-2 lóculos más de lo normal, que también contienen óvulos, pero solamente uno o dos lóculos son funcionales (foto 15).
Semilla Posee forma ovoide que se va adelgazando gradualmente hacia el pico. Presenta una depresión circular y un lomo, la quilla o carena de Bioletti, a ambos lados del cual hay dos surcos o fosetas.
Embrión La semilla de la vid es albuminada por lo que el embrión maduro ocupa una pequeña parte de la semilla junto al pico. El embrión es recto, con dos cotiledones, un corto hipocotilo, y el epicotilo.
Endosperma El endosperma de la vid es ruminado (con dos fosetas o increscencias ruminadas ambos lados del rafe), con forma irregular, formado por pequeñas células poligonales con pequeños espacios intercelulares. Contiene aceite, granos de aleurona e idioblastos.
Tegumentos El tegumento externo se alarga en la zona de la micrópila para formar el pico, con grupos de esclereidas e idioblastos. La capa más externa papirácea tiene una gruesa cutícula. La capa media es parenquimatosa o colapsada. La capa interna dura presenta esclereidas con pared secundaria lignificada, cada una con un cristal. El tegumento interno tiene su capa más externa con engrosamientos espiralados y la capa más interna, rica en taninos y de paredes radiales con engrosamientos irregulares.