Estimados lectores voy a copiar integro este articulo debido a la calidad del mismo podéis encontrarlo integro en el blog fitoagro.com
3. COMO ATACAN LOS PATÓGENOS A LAS PLANTAS
La superficie de una planta está compuesta por:
– Celulosa: células epidérmicas de la raíz y espacios intercelulares del parénquima de las hojas.
– Cutícula: recubre las paredes de las células epidérmicas de los órganos aéreos. Es frecuente que la cutícula de las partes jóvenes se recubran de una capa cerosa adicional.
Los patógenos para nutrirse necesitan penetrar las paredes externas de los órganos de las plantas para alcanzar los contenidos celulares que transforman en unidades más simples.
Las plantas reaccionan ante la presencia del patógeno produciendo estructuras y sustancias químicas que tratan de interferir el desarrollo el patógeno.
Para que un patógeno infecte a una planta debe ser capaz de abrirse pasa para alcanzar el interior de ésta, obtener los nutrientes y neutralizar las reacciones de defensa de ella.

3.1. FUERZAS MECÁNICAS QUE EJERCEN LOS PATÓGENOS SOBRE LOS TEJIDOS DE SUS HOSPEDANTES.
Parece ser que solo algunos hongos, planas superiores parásitas y nematodos aplican una presión de tipo mecánico sobre la superficie de la planta que intentan penetrar.
Antes de poder ejercer la presión para penetrar la superficie de una planta los patógenos necesitan adherirse a ella. Existen sustancias mucilaginosas que rodean a las hifas y radículas de plantas parásitas, pero la fijación parece estar gobernada por fuerzas intermoleculares que actúan al entrar en contacto las células del patógeno y del hospedador.
Una vez que se ha producido el contacto, en las hifas de los hongos y en la radícula de las plantas parásitas se una estructura aplanada y en forma de bulbo llamada apresorio; que permite una fijación más firme y que da lugar a un “punto de penetración” por el cual pasa una hifa (filamento) más delgado que ejerce la fuerza. El filamento aumenta su diámetro en cada penetración de la cutícula.
En el caso de los nematodos la penetración se hace mediante el estilete. Los labios se fijan a la superficie de la planta y mediante movimientos de su cuerpo hincan el estilete.
Toda vez que se ha producido la penetración el hongo o el nematodo secreta cantidades creciente de enzimas que ablandan o disuelven la pared celular.
3.2. DEGRADACIÓN ENZIMÁTICAS DE LAS SUSTANCIAS DE LA PARED CELULAR
Las paredes de las células epidérmicas suelen contener además, proteínas y lignina.
En el proceso de penetración la secuencia sería:
A.- Cera Cuticular: no se conoce, hasta la presente, ningún patógeno que produzca enzimas u otras sustancias degradativas de las ceras, por lo cual se supone que son penetradas de forma mecánica.
B.- Cutina: es la principal componente de la cubierta cuticular. La parte superior de esta cubierta se encuentra combinada con ceras; la inferior donde se une con las células epidérmicas) la péctina y la celulosa son fundamentales. La cutina es un poliéster insoluble.
Se sabe que muchos hongos producen cutinasas, enzimas que degradan la cutina. Las cutinasas son esterasas que rompen los enlaces éster de las moléculas cutina.
Parece que en el proceso, los Hongos producen continuamente pequeñas proporciones de cutinasa, que estimularía a las células de la epidermis a que la produjese también. Parece que las ceras existentes en la superficie de los hospedador también estimulan a los hongos a producir cutinasa.
La producción de cutinasa no tiene, por el momento, una relación directa con la patogeneicidad.
C.- Sustancias Pépticas: son el componente principal de la lámina media (es el cemento que mantiene a las células en su posición). Forman parte de las células de los tejidos en su pared celular primaria (forman un gel amorfo que llena los espacios que hay entre las microfibrillas de las células).
Las sustancias pépticas son polisacáridos que contiene, esencialmente, cadenas del ácido galacturónico. Las encargadas de degradar las sustancias pécticas son las pectinasas o enzimas pectolíticas.

– Péctina metilesterasas: cortan pequeñas porciones de las cadenas pécticas sin afectar sensiblemente su longitud, pero incrementan su solubilidad.
— Poligalacturonasas: rompen las cadenas pécticas al añadir una molécula de agua e hidrolizar el enlace entre dos moléculas de ácido poligalacturónico.
— Pectinaliasas o transeliminasas: rompen las cadenas de ácido poligalacturónico eliminando una molécula de agua del enlace, liberando cadenas pectínicas con un doble enlace insaturado.
Los patógenos pueden producir en todo momento pequeñas cantidades de enzimas pectinolíticas en presencia de pectinas que liberan monómeros o polímeros de ácido galacturónico. Si estos monómeros o polímeros son absorbidos por el patógeno funcionan como inductores de la síntesis y liberación de enzimas pectinolíticas (inducción autocatalítica). Cuando la concentración de monómeros o polímeros de ácido galacturónico es grande, ésta inhibe la síntesis de más enzimas (represión catalítica). La producción de enzimas es inhibida en presencia de glucosa.
— Las enzimas que degradan las pectinas participan en el desarrollo de muchas enfermedades, especialmente en las que se producen lapodredumbre blanca de los tejidos (maceración de los tejidos: ablandamiento pérdida de la cohesión de los tejidos, disgregación de las células que finalmente mueren).

D.- Celulosa: es un polisacárido formado por cadenas de moléculas de glucosa. En todas las plantas superiores constituye el armazón de las paredes celulares y se encuentra en forma de microfibrillas.
La degradación por enzimas conocidas, genéricamente, como celulasas da como resultado final la producción de moléculas de glucosa.
— La celulosa C1 rompe la celulosa nativa quebrando los enlaces.
— La celulosa C2 rompe la celulosa en cadenas cortas que son transformadas en celobiosa.
— La celobiosa es degradada a glucosa por el enzima -glucosidasa.
Se ha demostrado que las celulasas son producidas por varios hongos, bacterias y nematodos fitopatógenos.
Las enzimas celulolíticas ablandan los tejidos de las plantas vivas al tiempo que liberan azúcares (glucosa) que les sirven de alimento al patógeno.
E.- Hemicelulosa: mezclas complejas de polímeros de polisacáridos cuya composición y frecuencia parece variar dentro de los tejidos de la misma planta entre especies vegetales y según el desarrollo de la planta.
Son un componente de la pareces primaria y secundaria de la célula y forma parte, también, de la laminilla media.
— Las enzimas que intervienen en la degradación de la hemicelulosa se denominan, genéricamente, hemicelulasas. Según el monómero liberado del polímero sobre el que actúan, reciben nombres particulares: xilanasa, galactanasa, glucanasa, arabinasa, manasa, etc.
Al parecer las hemicelulasas son producidas por algunos hongos fitopatógenos, pero se desconocen como intervienen en la degradación de la pared celular del hospedante.
F.- Lignina: es un polímero amorfo tridimensional. Se encuentra en la laminilla media y en la pared celular de los vasos xilemáticos y de las células epidérmicas. Se ha señalado cierto número de especies de hongos como lo únicos que descomponen la madera. Las enzimas que secretan éstos se llaman ligninasas. Parece que algunos hongos y bacterias fitopatógenas producen ligninasas, aunque se desconoce si participan en la patogénesis
3.3. DEGRADACIÓN ENZIMÁTICA DE LAS SUSTANCIAS CONTENIDAS EN LAS CÉLULAS VEGETALES.
Los patógenos pueden, al parecer, absorber nutrientes formados por moléculas pequeñas (azúcares, aminoácidos). Sin embargo, algunos constituyentes del protoplasto celular deben ser degradados antes de ser consumidos (almidón, proteínas, grasas). Degradación que se produce mediante enzimas secretadas por el patógeno.
A.- Proteínas: los patógenos son capaces de degradar muchos tipos de proteínas. Las enzimas que utilizan son semejantes a las de los animales y plantas superiores. Genéricamente se las denomina proteinasas. Es componente estructural de las membranas de las células vegetales y actúan como enzimas.
B.- Almidón: las amilasas degradan el almidón a glucosa, directamente asimilable por el patógeno. Es el principal polisacárido de reserva que se encuentra en la célula vegetal.
C.- Lípidos: varios hongos, bacterias y nematodos tienen capacidad para degradar a los lípidos. Las enzimas reciben nombres específicos como lipasas, fosfolipasas, etc. los más importantes en la célula vegetal: grasas, aceites, fosfolípidos y glucolípodos (sirven de reserva, o junto a las proteínas componente esencial de las membranas celulares).

3.4. TÓXINAS MICROBIANAS. SU RELACIÓN CON LAS ENFERMEDADES DE LAS PLANTAS.
Las toxinas actúan directamente sobre los protoplastos vivos, otras actúan sobre el
protoplasma en general, las hay inhibidoras de enzimas, las hay que inhiben la permeabilidad de la membrana celular; otras, en fin, actúan como antimetabolitos que propician la deficiencia de un factor esencial para el desarrollo normal.
Los hongos y bacterias producen toxinas tanto en plantas hospedadoras como en medios de cultivo. Inducen alteraciones de reacciones metabólicas.
3.5. TÓXINAS QUE AFECTAN A UN AMPLIO NÚMERO DE PLANTAS HOSPEDANTES.
Se ha comprobado que la toxinas producidas por algunos fitopatógenos son capaces, por si mismas, de producir el síndrome total o parcial de la enfermedad sobre el hospedador al cual enferma el patógeno; y, también, a otros hospedadores a los cuales el parásito nunca enferma. Son toxinas no específicas que no son esenciales para que el microorganismo cause la enfermedad.
A.- Tabtoxina:
Se aisló y caracterizó por primera vez de la bacteria Pseudomonas syringae pv tabaci. Hoy se sabe que la producen otras cepas de patovar (pv) tabaci que enferman a soja y a la judía. Como tal no es tóxica, pero cuando se libera en la célula al hidrolizarse inactiva a la enzima glutamina sintetasa haciendo que la célula pierda su capacidad para defenderse frente a la bacteria.
B.- Faseolotoxina:
Está producida por la bacteria Pseudomonas syringae pv phaseolicola agente causal de la “grasa de la judía” y de otras leguminosas.
C.- Tentotoxina:
Está producida por el hongo Alternaria tenuis que causa clorosis en las plántulas de muchas especies, pudiendo originar muerte o en el mejor de los casos perdidas notables.
D.- Fusicoccina:
Está producida por el hongo Fusicoccum amygdali que produce la muerte de ramillas de almendros y melocotones.
E.- Otras toxinas no específicas del hospedante:
Producidas por Hongos:
-Ácido Fumárico, producido por Rhizopus spp agente causal de podredumbres en frutos en conservación.
– Ácido Oxálico, producido por Sclerotium que infecta a diversas especies.
– Ácido Alternárico, Alternariol y ziniol producidos por Alternaria spp que causa manchas foliares en numerosas especies.
– Ácido Fusárico producido por diversas formas especializadas del hongo Fusarium oxysporum.
Micotoxinas (introducidas por hongos): representan una amenaza para la salud del hombre y de los animales, debido a las dosis subagudas presentes en los animales y forraje. Principales: Atlatoxina (Aspergillus spp), toxinas de fusarium.
Producidas por Bacterias:
– Coronatina producida por Pseudomonas syringae pv atropurpurea que afecta a las gramíneas para pasto y a la soja.
– Siringomicina producida por Pseudomonas syringae pv syringae que causa manchas foliares en cítricos y otras plantas.
Toxinas específicas:
Son toxinas que solo actúan sobre hospedantes a los que enferma el patógeno que las produce y tienen poca o nula actividad sobre plantas no susceptibles. La mayoría de estas toxinas deben estar presentes para que el microorganismo sea capaz de producir enfermedad. Normalmente son producidas por hongos.
A.- Victorina o toxina HV:
Está producida por el hongo Helminthosporium victoriae. Enferma a las hojas basales de las plantas de avena susceptibles y produce una toxina que es llevada hasta las hojas superiores a las cuales seca, llegando a matar a las plantas. La toxina reproduce todos los síntomas externos de la enfermedad como si el patógeno estuviese presente. Solo las cepas del hongo que producen victorina son capaces de atacar a la avena.
B:- Toxina T:
Helminthosporium maydis produce una enfermedad foliar del maíz conocida como helmintosporiosis o tizón foliar. El hongo tiene una raza T, que no se distingue de las otras más que por su capacidad para producir la toxina T. Dicha raza solo ataca a las variedades de maíz que tienen citoplasma androestéril, la resistencia al hongo y a la toxina se hereda por vía materna.
C.- Toxina AK:
Producida por un patotipo de Alternaria alternata que causa la mancha negra en los perales japoneses. Los perales resistentes no manifiestan síntomas al ser pulverizados con la toxina AK, los sensibles si.
D.- Toxina AM:
Está producida por otro patotipo de Alternaria alternata que ataca a las hojas del manzano, originando manchas negras. Las resistentes toleran bien la toxina.
3.6. REGULADORES DEL CRECIMIENTO EN LAS ENFERMEDADES DE LAS PLANTAS.
El crecimiento de las plantas está bajo el control de un pequeño grupo de compuestos que en la naturaleza actúan como hormonas a los que se denominan reguladores del crecimiento:
Los más importantes: auxinas, giberelinas y citoquininas. Otros como el etileno
tienen un papel regulador en el desarrollo de las plantas.
El parásito ocasiona con frecuencia un desequilibrio en el sistema hormonal de la
planta y da lugar a síntomas anormales en aquella.
La acción del patógeno puede ser debida tanto a la secreción de reguladores del crecimiento en la planta infectada o mediante los efectos que producen sobre los sistemas reguladores de aquellos. Así, los achaparramientos, la emisión anormal de raicillas, alteraciones en la flor o frutos, el alargamiento de entrenudos, las epinastias, etc. son expresiones sintomatológicas de las alteraciones hormonales.
Auxinas:
La auxina presente de forma natural en las plantas es el ácido indolacético (IAA), que se produce de manera constante en los tejidos de las plantas en crecimiento y no se acumula ya que es degradada, siempre está en bajas proporciones en los tejidos sanos. Produce un aumento general en la respiración de los tejidos vegetales y promueve la síntesis del ARN mensajero y, en consecuencia, de las proteínas-enzimas y proteínas estructurales.
Muchas plantas infectadas por hongos, bacterias, nematodos, virus, micoplasmas muestran un aumento en los niveles de la auxina IAA.
— Hongos como Plasmodiophora brassicae, Phytophthora infestans, Fusarium oxysporum fsp cubense.
— Nematodos como Meloidogyne spp.
Inducen el incremento de IAA en sus hospedadores, aunque ellos pueden, también, producir dicha auxina.
El aumento del IAA parece ser debido a que el patógeno destruye la enzima IAA oxidasa.
Giberelinas:
Son constituyentes normales de las plantas verdes, aunque hay varios microorganismos que las producen. Fueron aisladas por primera vez a partir del hongo Gibberella fujikuori que causa la enfermedad conocida como “bakanae”del arroz. La giberelina más conocida es el ácido giberélico.
Las giberelinas aceleran el crecimiento de variedades enanas para que alcancen su tamaño normal. Favorecen la floración y el crecimiento del fruto. Se asemejan en su acción al IAA e inducen la formación de dicha hormona. Giberelinas y auxinas pueden actuar sinergísticamente.
Citoquininas:
Son potentes factores del crecimiento necesarios para la diferenciación y el crecimiento celular. Inhiben el envejecimiento al impedir la degradación de las proteínas y de los ácidos nucleicos. Se encuentran en bajas concentraciones en las partes verdes de las plantas, semillas y en la savia.
Su papel en enfermedades de las plantas no es bien conocido. Se sabe que su presencia aumenta en los tejidos afectados por roya donde ejercen una función de rejuvenecimiento en los tejidos afectados y de envejecimiento en el resto. También se sabe que su actividad disminuye en la savia y en los extractos de plantas de algodón infectadas con Verticillium dalia así como en plantas que sufren sequía.
Etileno (CH2=CH2):
Se produce de forma natural en las plantas y ejerce en ellas una serie de efectos: clorosis, abscisión de hojas, epinastia, maduración de frutos y aumento en la emisión de raíces adventicias. Posiblemente al inducir la formación de fitoalexinas u enzimas incremente la resistencia de la planta a patógenos.
Lo producen varios hongos y bacterias fitopatógenas.
Ácido Abcísico:
Es uno de los inhibidores del crecimiento que producen las plantas y algunos hongos.
Induce la dormancia, germinación de semillas, cierre estomático y estimulación de la germinación de las esporas de hongos.
La plantas enfermas lo tienen más que las sanas.
Polisacáridos
Hongos, bacterias y nematodos liberan constantemente cantidades variables de sustancias mucilaginosas que los protegen, cuyo papel está limitado a las enfermedades vasculares. Son liberados por los patógenos en los vasos xilemáticos obstruyéndolos, marchitamiento.
4. EFECTOS DE LOS PATÓGENOS SOBRE LA FISIOLOGÍA DE LAS PLANTAS.
4.1. EFECTO DE LOS PATÓGENOS SOBRE LA FOTOSINTESIS.
El procesos de la planta permite trasformar la energía luminosa en energía química que es utilizada internamente en las plantas para las actividadescelulares de la misma, cuando un patógeno ataca a la planta es evidente en cualquier alteración que esta presente puesto que la fotosíntesis genera la energía para el funcionamiento de la planta.Cuando observamos una planta que se encuentra en buen estado y al paso del tiempo la planta empieza a mostrar manchas coloriticas, lesiones necróticas y demás alteraciones visibles a simple vista , podemosconcluir que hay un ataque de un agente causal de enfermedad o patógeno.
4.2. EFECTO DE LOS PATÓGENOS SOBRE LA RESPIRACIÓN DE LA PLANTA HOSPEDANTE.
La respiración es el proceso mediante el cual las células, por medio de la oxidación (combustión) enzimáticamente controlada de los ácidos grasos y carbohidratos ricos en energía, liberan energía en una forma que puede ser utilizada para la ejecución de varios procesos celulares. Las células vegetales llevan a cabo la respiración básicamente en dos etapas. La primera implica la degradación de la glucosa hasta piruvato y se efectúa (ya sea en presencia o en ausencia de oxígeno), mediante la participación de las enzimas que se encuentran en el citoplasma de las células.
4.3. RESPIRACIÓN DE LAS PLANTAS ENFERMAS.
Cuando las plantas son infectadas por los patógenos, su tasa respiratoria por lo general 24 aumenta. Esto significa que los tejidos que son afectados utilizan con mayor rapidez su reserva de carbohidratos que los tejidos sanos. El aumento en la respiración se produce poco después de haberse producido la infección (casi en el momento en que aparecen los síntomas observables) y continúa aumentado durante la multiplicación y esporulación del patógeno. Después de esto, la respiración disminuye hasta su nivel normal e incluso hasta un nivel mucho menor que el de las plantas sanas. La respiración aumenta con mucha más rapidez en las infecciones de las variedades resistentes, en las que requieren y utilizan grandes cantidades de energía para activar rápidamente los mecanismos de defensa de las células del hospedante. Sin embargo, en dichas variedades, la respiración disminuye también con gran rapidez una vez que se ha llegado hasta un punto máximo. En tanto las variedades susceptibles, en las que los mecanismos de defensa no pueden activarse con rapidez ante el ataque de algún patógeno en particular, la respiración aumenta lentamente después de haberse producido la infección, pero continúa aumentando y se mantiene a un alto nivel durante mucho más tiempo.
4.4. EFECTO DE LOS PATÓGENOS SOBRE LA PERMEABILIDAD DE LAS MEMBRANAS CELULARES
Las membranas celulares consisten en una doble capa de moléculas lipídicas en las que están embebidas muchos tipos de moléculas proteínicas, parte de las cuales sobresalen, en general, a ambos lados de dicha bicapa. Las membranas funcionan como barreras de permeabilidad que permiten la entrada en la célula únicamente de sustancias útiles y evitan la salida de aquellos que le son necesarias. La bicapa lipídica es impermeable a la mayoría de las biomoléculas. Las pequeñas moléculas hidrosolubles como iones (átomos con carga eléctrica o electrólitos), azúcares y aminoácidos fluyen o son bombeados a través de canales especiales de membranas constituidos por proteínas. En las células vegetales, debido a la presencia de la pared celular, sólo moléculas pequeñas llegan a la membrana celular. Sin embargo, en las células animales y en los protoplastos vegetales preparados artificialmente, las macromoléculas o partículas pueden llegar también a la membrana celular y entrar en la célula por endocitosis; esto es parte de la membrana que rodea y forma una vesícula en torno al material que es llevado y liberado dentro de la célula. La disolución o perturbación de la membrana celular mediante factores químicos o físicos altera (o generalmente incrementa) su permeabilidad, y esto trae como consecuencia la pérdida incontrolable de sustancias útiles, así como también la posibilidad de inhibir la entrada en la célula de sustancias indeseables o de cantidades excesivas de todo tipo de sustancias.
5. COMO SE DEFIENDEN LAS PLANTAS DE LOS PATÓGENOS
Es impresionante la forma de cómo las plantas encuentran la forma de defenderse para evitar enfermarse y como las bacterias, hongos, virus, nematodos, micoplasmas, etc. Es verdad que las plantas sonafectadas en cada momento de su desarrollo son atacadas por cientos o miles de patógenos en ocasiones los patógenos son de la misma clase, la mayoría de las veces de que son atacadas les ocasionandaños considerables o menores la mayoría de los ataques llegan a sobrevivir en la cual continua con su ciclo normal.
5.1. DEFENSA ESTRUCTURAL.
La primera línea de defensa de las plantas ante el ataque de los patógenos es su superficie, la cual estos últimos deben penetrar para causar infección. Algunas defensas 25 estructurales se encuentran ya en las plantas incluso antes de que el patógeno entre en contacto con ellas. Esas es- tructuras incluyen la cantidad y la calidad de la cera y de la cutícula que cubren a las células epidérmicas, la estructura de las paredes celulares de estas últimas, el tamaño, localización y forma de los estomas y lenticelas y, por último, la presencia en la planta de tejidos protegidos por paredes celulares gruesas que obstaculizan el avance del patógeno.
5.2. ESTRUCTURAS DE DEFENSA QUE SE FORMAN EN RESPUESTAS A LAS INFECCIONES PATOGÉNICAS.
Aunque algunos patógenos se ven imposibilitados de entrar e invadir a las plantas debido a que éstas presentan estructuras de defensa internas o superficiales preformadas, la mayoría de ellos logran penetrar en sus hospedantes y producir diversos grados de infección. Sin embargo, incluso después de que el patógeno ha penetrado las estructuras de defensa preformadas, las plantas muestran varios grados de resistencia que consisten en la formación de uno o más tipos de estructuras que tienen más o menos una cierta efectividad para contrarrestar las invasiones posteriores de los patógenos sobre ellas.
5.3. ESTRUCTURAS HISTOLÓGICAS DE DEFENSA.
Formación de capas de corcho La infección que ocasionan los bongos, bacterias e incluso algunos virus y nematodos en las plantas con frecuencia induce la formación de varias capas de células de corcho más allá de la zona donde se ha producido la infección (figuras 5-1 y 5-2), aparentemente como resultado de la estimulación que ejercen las sustancias que secreta el patógeno sobre las células del hospedante. Las capas de corcho no solo inhiben el avance que pueda intentar el patógeno sobre las otras células, sino también bloquean la difusión de cualquier sustancia que el patógeno pueda secretar. Además, estas capas detienen el flujo de agua y nutrientes desde las zonas sanas hasta la zona infectada y suprimen la nutrición del patógeno.
5.4. FORMACIÓN DE LAS CAPAS DE ABSICIÓN.
Las capas de abcisión se forman sobre las hojas jóvenes y funcionales de los árboles frutales de hueso duro después de haber sido infectadas por cualquiera de los distintos hongos, bacterias o virus (véase la figura 12-11). La capa de abscisión consta de una abertura que se forma entre dos capas circulares de células foliares en torno al foco de infección. Después de que ésta se ha producido, la lámina media localizada entre esas dos capas de células se disuelve a lo largo del grosor de la hoja, separando por completo la zona central del resto de la hoja (figura 5-3). Gradualmente, esta zona se seca, muere y desprende, llevándose consigo al patógeno. De esta forma, la planta, al deshacerse de esa zona infectada junto con unas cuantas células todavía sanas, impide que el resto de los tejidos de la hoja sean invadidos por el patógeno o que sean afectados por sus secreciones tóxicas.
5.5. FORMACIÓN DE TÍLIDES.
Las tílides se forman en los vasos xilemáticos de la mayoría de las plantas que se encuentran bajo condiciones ambientales adversas y durante la invasión que llevan a cabo la mayoría de los patógenos vasculares. Las tílides son crecimientos excesivos del protoplasto de las células parenquimatosas adyacentes, las cuales se proyectan hacia los vasos xilemáticos a través de puntuaciones (figura 5-4). Las tílides presentan paredes celulósicas y pueden, debido a su tamaño y abundancia, obstruir completamente a los vasos. En algunas variedades de planta, las tílides se forman rápida y abundantemente frente al patógeno (en tanto éste se encuentre en las raíces jóvenes), bloqueando su avance y por lo tanto, permitiendo que las plantas permanezcan Ubres y sean resistentes a ese patógeno. Las variedades vegetales que forman pocas tílides o ninguna frente al patógeno son siempre susceptibles a las enfermedades.
5.6. REACCIÓN DE DEFENSA NECRÓTICA.
Defensa mediante hipersensibilidad En la mayoría de las relaciones que se establecen entre el hospedante y el patógeno, este último puede llegar a penetrar la pared celular, pero tan pronto como entra en contacto con el pro- toplasto de la célula, el núcleo se desplaza hacia él y en poco tiempo se desintegra y aparecen en el citoplasma granulos de color café y en forma de resina, primero en torno al patógeno y después por todo el citoplasma. Mientras aumenta el cambio de coloración del citoplasma de la célula vegetal y se produce la muerte, las 29 hifas del hongo empiezan a degenerar (figura 5-7). En la mayoría de los casos, las hifas no se desarrollan fuera de las células vegetales, por lo que cesa la invasión de esas hifas.
6. GENÉTICA DE LAS ENFERMEDADES DE LAS PLANTAS.
La producción agrícola vegetal en el mundo incluye rubros prioritarios como los cereales, las raíces y tubérculos, leguminosas de grano, oleaginosas, frutas y hortalizas, de amplio consumo. El rendimiento de cada uno de estos rubros depende de la eficiencia en el uso de la tierra, del capital y mano de obra, pero sobre todo de la aplicación de tecnologías limpias, de nuevas semillas y plantas resistentes a condiciones adversas y manejo adecuado de condiciones ambientales, económicas, políticas, vinculados a la actividad agrícola. La importancia de conocer aspectos relacionados con la información genética de los microorganismos, el control de las enfermedades a nivel cultural, biológico y genética hace del futuro profesional una persona fundamental en el tratamiento eficiente de los problemas sanitarios en la agricultura.
6.1. INFORMACIÓN GENÉTICA.
La información genética de los microorganismos se encuentra codificada en el Acido ácido DNA, presente en los cromosomas y en el caso de los virus en el ácido RNA. Los patógenos que infectan a los cultivos generalmente son específicos para causar la enfermedad en esa planta y no en otra, como es el caso elFusarium oxysporum que afecta al tomate y no a otra planta, existen otros tipos de Fusarium exclusivos para ocasionar enfermedades en muchos cultivos, dicha especificidad es proporcionada por genes que actúan sobre ese hospedante en particular , de manera que según afirma Agrios G (2001 ) cada hospedante tiene un juego de genes que determina su susceptibilidad contra un patógeno y son a su vez específicos para el patógeno.
Algunos microorganismos afectan a muchas especies de plantas por que contienen genes para desarrollar virulencia más amplia sobre diversos hospedantes, en esa misma dirección el hospedante también posee genes que le permiten ser susceptible a la virulencia de cientos de patógenos. Estos genes aparecen por mutación tanto en patógenos como hospedantes y producen resistencia a la infección.
6.2. MECANISMOS DE VARIABILIDAD.
¿Cuales son los mecanismos de variabilidad?
Recombinacion: este proceso comienza con la reproducción de tipo sexual de hongos y nematodos, en donde dos núcleos haploides con material genético distinto, se unen para conformar un núcleo diploide o cigoto. Se presenta entrecruzamiento genético en las cromátidas de un cromosoma con partes de las cromatidas del otro cromosoma. En los hongos se producen hifas que forman micelio y esporas haploides
Mutaciones: consiste en un cambio en el material genético de un organismo transmitido hereditariamente a la progenie.
La mutación consiste básicamente en un cambio en la secuencia de bases DNA por adición o sustitución de bases. En algunas ocasiones ocurren en las patógenas mutaciones extracromosomicas y en ese caso los organismos adquieren capacidad para desarrollar algunos procesos fisiológicos no comunes en ellos.
6.3. MECANISMOS ESPECIALIZADOS DE VARIABILIDAD.
Parasexualismo: se dan recombianaciones geneticas en los heterocariones de los hongos, se dan entrecruzamientos geneticos en las divisiones micóticas.
Heterocariosis: se produce a través de procesos de fertilización y anastomosis.
Heteroploidia: se da en organismos con un número de cromosomas por núcleo diferente al normal 1N o 2N. La heteroploidía afecta el número de esporas, el color de las hifas y del micelio.
Recombinación genética: En los virus inoculados en un mismo hospedante, y que provienen de una misma cepa, al reaislar se produce un virus con características disimiles a los originalmente inoculados en el hospedante.
6.4. RESISTENCIA DE LAS PLANTAS A LAS ENFERMEDADES.
Resistencia verdadera: es la resistencia a enfermedades controlada por uno o varios genes. El patógeno y el hospedante son incompatibles y no hay reconocimiento químico entre ellos.
Resistencia Horizontal se denomina poligénica o múltiple cuando la resistencia está a cargo de muchos genes, este tipo de resistencia retarda el desarrollo de las enfermedades y retrasa la propagación de enfermedades.
Resistencia vertical se presenta en plantas que son resistentes a algunas razas de patógenos y susceptibles a otras razas del mismo patógeno. se denomina resistencia monogénica u olegogénica controlada por uno o varios genes, el hospedante por lo general responde reaccionando a la virulencia; algunas especies tienen 20 o 40 genes resistentes contra un patógeno
Resistencia aparente: Se traduce en la tolerancia de las plantas a una enfermedad.
6.5. RECOMBINACIONES GENÉTICAS.
Genes de resistencia
Genes recesivos/Susceptibilidad
genes virulentos/genes no virulentos
R dominante ( resistente )
r recesivo ( Susceptible )
A Dominante ( No virulento )
AR (-)
Ar( + )
a recesivo ( virulento )
aR (+)
ar (+)
De las combinaciones mostradas en la tabla anterior, la AR es resistente; en la Ar el hospedante carece de resistencia, luego es infectado; en aR el hospedante es infectado y en ar la planta es infectada debido a que es virulento y susceptible. Existen genes estructurales que determinan la clase y propiedades de las proteínas, los genes reguladores controlan el ritmo de actividad de los genes y la síntesis de proteína
Resistencia de las plantas: Las técnicas de mejoramiento genético para obtener resistencia se describen a continuación:
Fusion de protoplastos: consiste en la fusión de células híbridas que contienen cromosomas y citoplasma de plantas emparentadas e incorporan nuevas características a partir de la fusión de protoplastos, así se regeneran nuevas plantas con mayor número de cromosomas y nuevas características a partir de la fusión.
Transformación genética de células resistentes a enfermedades: Consiste en la introducción de DNA en los cromoplastos de las células bien sea por absorción directa del DNA, inyección del DNA o transferencia de DNA o lisosomas y posterior transferencia de la molecular que lleva la información mediante los plasmodios del centrómero… El Agrobacterium se ha empleado para introducir el las plantas nuevos genes. El estudio lo Reporta Agrios G (2001) cuando refiere investigaciones sobre aislamientos del gen de las proteínas de la semilla del frijol y al introducirlo en el plasmodio de agrobacterium, para generar tumores. Cuando la enfermedad se produce una mínima parte del DNA del plasmodio Ti o inductor de tumores, se transfiere a la célula vegetal y lo incorpora dentro del genoma de la planta. La resistencia de la planta se expresa al incorporar genes de resistencia de manera que puedan actuar en el caso de que se presente una enfermedad, la planta puede generar resistencia al ataque de los patógenos.
Existen técnicas de resistencia inducida por microbios como una estrategia novedosa para el control de enfermedades en las hortalizas sobre todo las transmitidas por insectos, según Geoffrey, Z (1996) las enfermedades de la plantas causados por patógenos que son a su vez transmitidos por insectos es uno de los problemas mas importante para las hortalizas, ya que son insectos de alta movilidad y colonizan rápidamente los cultivos. Enfermedades epidémicas como el CMV en tomate que produce el virus, tiene como vector un aphido. Las plantas han evolucionado hasta desarrollar mecanismos de defensa contra enfermedades. Kloepper (1982) acuño el término RSI o proceso de protección activa o sistémica de una planta, dependiendo del tipo de barreras físicas y químicas levantadas por las plantas. Ciertas Bacterias que colonizan las raíces de las plantas o rhizobacterias, que promueven el crecimiento de las plantas, controlan también algunos patógenos del suelo y funcionan como antagónicas de los patógenos del suelo mediante competencia. Algunos experimentos producidos en plantas reproducidas por semillas y tratadas con RPCP o bacterias promotoras del crecimiento (rhizobacterias), incidieron en la baja aparición de enfermedades como la marchites bacteria
También existen estudios en la evaluación de 26 lineas de RPCP o rhizobacterias realizados en Auburn en variedades de tomate, las semillas de tomate fueron mezcladas con granulados bacteriales antes de ser sembradas, las plantas fueron inoculadas con CMV o virus de tabaco por fricción una semana con rhizobacterias después del trasplante. El porcentaje de plantas con síntomas de virosis en los tratamientos fue de 14.7% en comparación con el 75.8 % de las plantas que no fueron tratadas con RPCP.
6.6. LA NATURALEZA DE LA RESISTENCIA A LAS ENFERMEDADES.
Desde que el hombre empezó su lucha contra las enfermedades de las plantas, se ha presentado el problema de por qué algunas plantas permanecen total o casi totalmente inalteradas por la enfermedad en tanto que otras no pueden soportarla, y todavía se busca la solución de este problema sin alcanzar la respuesta definitiva, lo cual no implica, necesariamente que ignoremos todos los fenómenos de la resistencia a la enfermedad, ya que sabemos que ésta es tan variable como es de esperarse en una rdación tan compleja como la que existe entre el susceptivo y el patógeno. El conocimiento. de los factores responsables de la inmunidad o de la resistencia a las enfermedades en las plantas, es indispensable para el ingeniero agrónomo, especialmente para los fitopatólogos, genetistas y fitotécnicos por cuanto sobre ese conocimiento se basa fundamentalmente uno de los métodos de control de mayores perspectivas: el de la Inmunización. Si bien es cierto que en los últimos años han aparecido muy interesantes artículos sobre este tema, la mayoría de la literatura está dispersa en publicaciones inaccesibles para muchos interesados y por este motivo el autor ha creído útil hacer un breve recuento informativo de las distintas teorías y hechos expuestos por los investigadores, al respecto, -sin que haya pretendido agotar el tema ya que ello sería imposible- para beneficio principalmente, de sus discípulos.
6.7. DEFENSA MECÁNICA
Cutícula.-Sin duda, el espesor de la cutícula desempeña un papel muy importante en la resistencia que ciertos susceptivos presentan a un patógeno. Sorauer, (citado por Appel, 1915: 780), encontró que la resistencia de ciertas variedades de claveles a la infección de la roya se debía al espesor de la cutícula.
El sugiere también la posibilidad de que este hecho se deba a la presencia de una capa cerosa que siempre está presente en las gramíneas.
Se ha sugerido que existe íntima relación entre el espesor del pellejo y la resistencia a la «pudrición Párda» en algunas variedades de ciruelas. Se ha encontrado que variedades de pellejo muy delgado compuesto de dos capas, además de la epidermis y con pulpa tierna, son más susceptibles a la enfermedad que las variedades de piel gruesa constituída por 5 a 8 capas de cé lulas y de una textura más firme y dura.
La resistencia de las plántulas de cereales a los carbones muestra la influencia de la cutícula sobre la infección causada por el patógeno; esta influencia obra solamente durante los primeros estados del desarrol’lo de la planta y se cree que es debida a la existencia de ácido silícico en los tejidos de la plántula (Appel 1915: 780).
Epidermis.-La naturaleza de las células epidermales del trigo se considera corno un factor de resistencia a la infección causada por las royas; las variedades Kota por ejemplo. tienen células epidermales muy duras lo cual exige para abrirse paso a través de ellas, una presión mayor de las hifas, con lo cual las pústulas se demoran más para presentarse y el período de infección se alarga. En otros casos las pústulas traspasan imperfectamente la epidermis o no alcanzan a sobrepasarla, permaneciendo por tanto en una condición subepidermal. (Stakman & Hart 1933: 259).
Estomas.-EI arreglo o disposición de los estomas, lo mismo que su forma y estructura, pueden ser de importancia en la klendusidad puesto que hay muchos casos evidentes en que ellos aparecen corno factores de primera importancia para evitar la entrada del patógeno.
6.8. PRODUCCIÓN DE VARIEDADES RESISTENTES.
El mejoramiento gen ético es un proceso mediante el cual son seleccionadas características de interés agronómico, transfiriéndose de un organismo a otro los genes que las codifican. Se logra mediante una serie de cruzamientos controlados, seguido de un proceso de evaluación y selección. La estrategia del programa de mejoramiento en café lo constituye la formación de líneas puras. Estas líneas se obtienen luego de una serie de procesos de autofecundación.
6.9. MEJORAMIENTO GENÉTICO DE PLANTAS PARA OBTENER RESISTENCIA A LAS ENFERMEDADES.
El mejoramiento de la resistencia a las enfermedades data de principios de este siglo, sin embargo mucho antes el hombre había observado y reportado un comportamiento diferente de las plantas a las enfermedades.Dos echos importantes marcaron el desarrollo del mejoramiento para la resistencia a enfermedades; uno la aceptación de la naturaleza parasítica de las enfermedades y el desarrollo de la genética como ciencia. El trabajo de la resistencia genética en la interacción huésped patógeno enmarca bases genéticas, fitopatológicas, fisiológicas, bioquímicas y moleculares.
El uso exclusivo de híbridos o variedades que son los más productivos, puede provocar que, en una región o en todo un país desaparezca la mayoría de las variedades nativas y con ellas las posibilidades futuras de mejorar las plantas cultivadas. Así, cuando en una región o país se agota la variabilidad genética, es necesario introducir nuevos materiales de la especie, de otros lugares donde exista gran variabilidad. Una vez que se han obtenido las colecciones de semillas de diferentes variedades, hay que clasificarlas, evaluarlas y conservarlas como semillas vivas, lo cual implica que todas ellas se siembren periódicamente, a fin de obtener semilla nueva (rejuvenecer).
La renovación de la semilla para su preservación ocasiona altos costos, debido a que:
No todas las colecciones pueden sembrarse en una misma localidad por problemas de adaptación a diferentes climas.
Es difícil manejar varios miles de lotes de diferentes plantas polinizadas artificialmente (plantas alógamas).
Es necesario manejar las colecciones muy cuidadosamente, a fin de evitar mezclas mecánicas, errores en las anotaciones o pérdidas por enfermedades o mal manejo de los materiales.
La viabilidad de la semilla depende fundamentalmente de la temperatura y la humedad. La mayoría de las semillas conservan su viabilidad por más tiempo cuando su contenido de humedad es muy bajo (4 a 7%), en una atmósfera seca o al vació (semillas enlatadas) y a una temperatura baja (de 2º a 5 ºC). Por lo tanto, la semilla se guarda a baja humedad en frascos de vidrio con insecticida, en cuartos refrigerados y aire seco. En estas condiciones, las semillas pueden sembrarse una vez cada diez años. Como el número de colecciones es muy grande, en vez de sembrarse todas ellas en una sola ocasión, lo recomendable es sembrarlas en grupos escalonados, de modo que cuando se siembre el último de los grupos, se vuelva a sembrar el primero y así sucesivamente.
7. EFECTO DEL AMBIENTE EN LA PRODUCCIÓN DE ENFERMEDADES INFECCIOSAS.
Los países invierten grandes cantidades de dinero para erradicar y combatir las enfermedades y las plagas de los animales y las plantas. El cambio climático está creando condiciones favorables para que se produzcan plagas y enfermedades de las plantas y los animales en nuevas regiones, y también está transformando sus vías de transmisión.
Si bien es evidente que el cambio climático está modificando la distribución de las plagas y las enfermedades de los animales y las plantas, es difícil prever todos los efectos de este cambio. La modificación de las temperaturas, la humedad y los gases de la atmósfera puede propiciar el crecimiento y la capacidad con que se generan las plantas, los hongos y los insectos, alterando la interacción entre las plagas, sus enemigos naturales y sus huéspedes. Las transformaciones que experimenta la cubierta vegetal de la Tierra, como la deforestación y la desertificación, pueden incrementar la vulnerabilidad de las plantas y los animales que quedan ante las plagas y las enfermedades.
7.1 EFECTO DE LA TEMPERATURA Y HUMEDAD
La supervivencia y función de la mayoría de los hongos fitopatógenos depende ampliamente de las condiciones predominantes de temperatura y humedad o de la presencia de agua en su medio. Un micelio libre sólo sobrevive dentro de un cierto intervalo de temperatura (que va de -5 a + 45°C) y cuando entra en contacto con superficies húmedas, ya sea que se localicen en el exterior o el interior de una planta hospedante., la mayoría de las esporas resisten intervalos bastante amplios de temperatura y humedad, y permite que el hongo sobreviva a los días cálidos y a las bajas temperaturas. Sin embargo, las esporas de los hongos requieren también humedad y temperaturas adecuadas para poder germinar.
7.2. EFECTO DEL VIENTO.
El viento ayuda a la propagación de algunas plagas y enfermedades (p.ej.: nubes de langostas, pulgones, escarabajo de la patata, oidio, mildiu, etc.); puede transportar semillas de malas hierbas; y, puede entorpecer la conservación de la pureza varietal en el proceso de producción de semilla.
8. ENFERMEDADES DE LAS PLANTAS CAUSADAS POR HONGOS.
La mayoría de los patógenos de plantas son hongos de las divisiones ascomycetes, basidiomycetes u oomycota.
Una enfermedad fúngica puede ser descrita como policíclica si el agente causal es capaz de producir esporas y reinfectar plantas durante una temporada de crecimiento, o monocíclica si el agente causal debe esperar una nueva temporada. Esta clasificación aplica a regiones con cuatro estaciones donde los patógenos deben producir estructuras específicas para sobrevivir al invierno. Los hongos fitopatógenos también pueden ser definidos por los tipos de esporas que producen y por el método por el cual penetran en la planta. Una vez que el patógeno ha penetrado produce un haustorio y crece dentro de la planta (alimentación biotrófica), o mata las células de su alrededor y se alimenta del tejido muerto (alimentación necrotrófica). La identificación de los patógenos se realiza basándose en los signos y síntomas de la enfermedad. Signos se refieren a la observación de alguna de las estructuras del patógeno (como esporulación). Síntomas son evidencia secundaria producida por la planta de que un patógeno esta presente (como el marchitarse de las hojas).
Las principales enfermedades causadas por hongos son mildius, oidios, royas, carbones, agallas y deformaciones, necrosis, chancros, marchiteces foliares, vasculares, etc., podredumbres radiculares, de flores, de frutos, etc. y micosis post-recolección, etc.
8.1. CARACTERÍSTICAS DE LOS HONGOS FITOPATÓGENOS.
La reproducción puede ser asexual o sexual, principalmente por esporas.
- La mayor parte de los hongos son saprofitos; algunos se consideran parásitos; otros mutualistas (que se asocian con otros seres en beneficio mutuo, como ocurre con los líquenes! Eucariontes (con pared celular)! Sin tejido vascular!
- Se reproducen principalmente por medio de esporas, que son diseminadas por el viento y pueden ser sexuadas o asexuadas.
- ! No tienen movimiento!
- Pueden ser uni o multicelulares!
- La pared celular esta formada de quitina; en las plantas es celulosa.! Son heterotróficos, se alimentan de materia orgánica. Los organismos autotróficos, producen sus alimentos por medio de la fotosíntesis! Los animales, también heterotróficos, primero ingieren y después digieren.
- Los hongos primero digieren y después ingieren!
- Para lograr lo anterior producen toxinas.!
- La mayoría de los hongos, almacenan alimentos como glicógeno, igual que los animales: mientras las plantas almacenan polisacáridos.!
- La mayoría de los hongos tienen un núcleo pequeño con poco DNA.!
- Son saprofitos! Se les considera parásitos y mutualistas.!
- Se alimentan de materia orgánica en descomposición.
- La falta de clorofila afecta profundamente su forma de vida: no necesitan de la luz
- crecen en cualquier dirección; invaden el sustrato con filamentos absorbentes.
- En los ecosistemas su función equivale alas aves de rapiña o los animales carroñeros.! Junto con las bacterias, su función principales reciclar el carbono, el nitrógeno y los minerales esenciales para la nutrición.!
- Como parásitos, usan la materia orgánica de organismos vivos, causando algún daño a plantas, animales y humanos.!
- En simbiosis, pueden beneficiar a otros organismos, como las mico rizas en las raíces de las plantas y los líquenes: asociación de un hongo y una alga.
- ! Son organismos muy útiles por su versatilidad genética y fisiológica.!
- Producen enormes cantidades de esporas que permanecen viables hasta que las condiciones climatológicas favorecen su multiplicación.!
- Pueden causar daños a los humanos por las toxinas que producen.!
- Descomponen toda clase de productos manufacturados, exceptuando los plásticos y algunos plaguicidas. Algunos como las levaduras, son muy útiles al ser humano. Los hay que producen antibióticos, hormonas y esteroides.!
- Son muy importantes en la investigación, debido a que se reproducen rápida y fácilmente, ocupan poco espacio, se multiplican rápidamente y su ciclo de vida es corto.
8.2. REPRODUCCIÓN DE LOS HONGOS FITOPATÓGENOS.
La reproducción en los hongos puede ser asexual o sexual y, en ambos casos, las esporas son las estructuras, responsables de dispersar la progenie para colonizar nuevas localizaciones. Algunas esporas están diseñadas para resistir condiciones adversas de crecimiento o para proporcionar un periodo de latencia. El micelio de los mohos puede también estar fragmentado, y los fragmentos resultantes pueden cada uno subsecuentemente desarrollarse en un talo individual por el proceso de reproducción vegetativa. El término reproducción vegetativa es usado para referirse a la reproducción asexual, donde las estructuras reproductoras especiales no son esporas.
8.3. REPRODUCCIÓN ASEXUAL DE LOS HONGOS FITOPATÓGENOS.
Reproducción asexual en hongos. La forma simple de la reproducción asexual es la producción de esporas vegetativas. Son dos estructuras principales, asociadas con la reproducción vegetativa. Son los artroconidios, y los clamidoconidios. Los artroconidios son producidos por las hifas que se separan y fragmentan. Estos pueden también ser denominados talosporas. Los clamidoconidios son usualmente mayores que los artroconidios, son redondeados y están inflados con alimentos de reserva. La formación de estas estructuras es usualmente como respuesta al estrés medioambiental. Bajo condiciones favorables, ambos, artroconidios y clamidoconidios germinan para producir un nuevo micelio. Las verdaderas esporas asexuales de los hongos difieren de las esporas vegetativas en que son formadas en estructuras especializadas llamadas esporóforos. Los cuales son también producidos generalmente en gran número. Se producen porque las células se dividen mitóticamente, y la composición genética de las esporas es idéntica a la de la célula parental.
Estas esporas son variables en tamaño, forma complejidad y color. Esto proporciona medios excelentes para la identificación de los hongos. Y también en parte forma la base de la clasificación micológica. Algunos hongos producen sólo un tipo de esporas asexuales, mientras otros pueden producir diferentes tipos de esporas. La mayoría de las esporas son diseminadas por el viento, agua o los insectos. Las esporas asexuales proporcionan un medio de reproducción para los hongos y, a causa del gran número de esporas que son producidas, la diseminación extensa de las especies es posible. Algunas esporas asexuales tienen vida corta, y son sensibles al estrés externo como la radiación ultravioleta y la desecación.
Sin embargo, este no es el caso siempre. Muchas esporas, particularmente aquellas que tienen pigmentación oscura o aquellas con pared gruesa, tienen resistencia a las presiones medioambientales. Las esporas resistentes pueden ser usadas como un estado de latencia en el ciclo de vida del hongo. Las esporas asexuales pueden ser producidas bien exógenamente en los extremos o lados de las hifas, o endógenamente en estructuras especializadas semejantes a sacos llamadas esporangios. Los hongos inferiores producen esporas en esporangios que están formados típicamente en los extremos de hifas ordinarias o en hifas especializadas denominadas esporangióforos, aunque pueden ser formados a lo largo de la estructura hifal.
Las esporas producidas exógenamente están formadas muchas veces en hifas especializadas, y son denominadas conidiosporas o simplemente conidias. Las conidias varían en forma, color y complejidad y pueden ser grandes o pequeños. Estas están unidas en estructuras llamadas conidióforos. Los conidióforos algunas veces, pero no siempre, difieren de las hifas vegetativas, y pueden ser característicos de un género o especie de hongo particular. Ver figura 1, reproducción asexual en hongos deuteromycetes.
8.4. REPRODUCCIÓN SEXUAL DE LOS HONGOS FITOPATÓGENOS.
Reproducción sexual en los hongos. Los hongos son clasificados de acuerdo a su método de reproducción sexual, pero hay un grupo de hongos en el que la reproducción sexual no ha sido observada y existen sólo en el estado anamórfico. Estos hongos son agrupados juntos en un grupo conocido los Hongos Imperfectos o Deuteromycetes.
La reproducción sexual en hongos es muchas veces en respuesta al estrés ambiental como cambios de temperatura, pH adverso o carencia de nutrientes. En el laboratorio, las condiciones pueden ser manipuladas para disparar la reproducción sexual por cultivos de crecimientos en medios que son deliberadamente bajos en nutrientes. El resultado de la reproducción sexual en hongos es la producción de esporas sexuales que son muchas veces estructuras resistentes, capaces de entrar en una fase de latencia. Como en toda reproducción sexual, en los hongos básicamente implica la fusión de dos núcleos compatibles. Con la excepción de algunas especies de levaduras, los hongos están en un estado haploide. Esto consiste en que tienen un solo conjunto de cromosomas desapareados.
La reproducción sexual de los hongos se produce un estado diploide, en el que los cromosomas están pareados, y la fusión celular es seguida por una meiosis del núcleo del zigoto (inmediatamente a menudo) así que la progenie puede regresar otra vez más al estado haploide. Típicamente ellos tienen tres fases en la reproducción sexual de los hongos, nombrada plasmogamia, cariogamia y meiosis. La plasmogamia supone la fusión de dos protoplastos. Esto conduce a unir dos núcleos compatibles en células idénticas.
El par de núcleos es llamado dicarion y la célula que los contiene se designa por dicariótica. En los hongos inferiores, la plasmogamia es casi inmediatamente seguida por la cariogamia, o la fusión de los dos núcleos, pero en los hongos superiores estos dos procesos pueden ser separados en el tiempo. Además, en los hongos superiores, las células dicarióticas pueden multiplicarse, con una división simultánea de los dos núcleos en cada célula. Este proceso puede producir numerosas células dicarióticas. Esto es lo que se refiere a la fase dicariótica. Cuando la fusión nuclear o la cariogamia sucede eventualmente, esta es seguida por la meiosis, volviendo las células fungales al estado haploide una vez más. Los órganos sexuales de los hongos se denominan gametangios, estos son diferenciados desde las hifas vegetativas.
En algunos hongos son indistinguibles unos de los otros, además otros gametangios machos y hembras son claramente diferentes.
8.5. REPRODUCCIÓN SEXUAL EN LOS FICOMICETOS.
En todos los Ficomicetos, el resultado de la reproducción sexual es producir esporas que germinan bajo condiciones favorables para producir el estado reproductivo asexual directamente, o poco después de la germinación. Los Ficomicetos acuáticos generalmente forman gametos móviles llamados zoosporas. Se fusionan para formar un zigoto móvil que solamente le queda entrar en una última fase. Algunos Ficomicetos tienen gametangios morfológicamente dispares. Las estructuras masculinas son llamadas anteridios, y las estructuras femeninas son oogonias. El zigoto que es producido por un anteridio y una oogonia es llamado oospora. En los ficomicetos más avanzados, los gametangios son hifas modificadas, morfológicamente. Estas fusiones terminan con la producción de paredes gruesas, estructuras resistentes llamadas zigosporas u oosporas.
8.6. REPRODUCCIÓN SEXUAL ES ASCOMICETOS.
La reproducción sexual en los Ascomicetos termina en la producción de ascosporas sexuales haploides en una estructura parecida a un saco llamada asca. La forma de las ascosporas y ascas varia con la especie particular de hongo. Típicamente hay 8 ascosporas en cada asca.
En los Ascomicetos inferiores, incluyendo las levaduras, el proceso de reproducción sexual es simple. Las dos células vegetativas se fusionan, y esto es seguido inmediatamente por la fusión de los dos núcleos. La célula zigoto resultante favorece al asca en la cual sucede la meiosis para producir las ascosporas. En los Ascomicetos superiores, la reproducción sexual es un proceso más complicado. Los núcleos compatibles han sido frecuentemente encontrados morfológicamente dispares en sus gametangios. El gametangio macho es llamado anteridio, y la hembra es ascogonio. La fusión nuclear, o cariogamia, no se hace inmediatamente a continuación de la plasmogamia, o fusión protoplástica, y las células dicarióticas son producidas. También son referidas como hifas ascógenas. Las ascas que contienen ascosporas son producidas desde las células binucleares terminales de las hifas ascógenas, y es aquí donde la cariogamia y la meiosis toman sitio eventualmente para producir las ascosporas. Son varios tipos de ascas. Algunos son globulares, en forma de porra, liberando sus ascosporas por ruptura de la pared, mientras otras son cilíndricas y tienen mecanismos de eyección de las esporas.
Los Ascomicetos superiores producen sus ascas en una estructura fructífera o ascocarpo (Ascoma). Hay tres tipos generales de ascocarpos, llamados cleistotecio, peritecio y apotecio. El cleistotecio tiene completamente cerrada su estructura donde encierra las ascas que dispone aleatoriamente dentro; las ascosporas pueden ser sólo liberadas por ruptura o desintegración de la pared. El peritecio tiene un estrato de ascas organizadas en sus paredes interiores, y un ostiolo a través del cual las ascosporas son liberadas. En muchas de Ascomicetos que producen peritecios, cada asca asciende al interior del ostiolo en sucesión., y allí descarga enérgicamente cada una de sus ascosporas. Un apotecio es una estructura abierta con ascas ordenadas en un estrato bien definido en su superficie superior. Cleistotecio y peritecio son producidos en cultivos, y son muchas veces visibles a simple vista.
8.7. REPRODUCCIÓN SEXUAL EN BASIDIOMICETOS.
La célula sexual característica en este grupo de hongos es el basidio. El basidio soporta las células sexuales haploides o basidiosporas, usualmente cuatro, externamente en estructuras llamadas esterigmas. En los Basidiomicetos inferiores el basidio está septado y produce desde una hifa o una pared engrosada la última espora llamada teliospora.
En los Basidiomicetos superiores los basidios son unicelulares y con forma de porra y son producidos generalmente en estructuras fructíferas conspicuas llamadas basidiocarpos. Muchas veces están organizadas en capas bien definidas llamadas himenio. Una fase micelial extensiva binucleada (dicariótica) es característica de estos grupos de hongos. Resulta de la fusión de hifas adyacentes, sin fusionar sus núcleos; éstas pueden ser de dos líneas sexuales compatibles si el hongo es heterotálico. Un proceso llamado formación abrazadera asegura que cuando estas células dicarióticas se multiplican, y sus dos núcleos se dividen simultáneamente, uno de cada par va dentro de cada célula hija.
8.8. NUTRICIÓN DE LOS HONGOS FITOPATÓGENOS.
Los hongos son muchas veces erróneamente confundidos con las plantas. Sin embargo las plantas pueden elaborar compuestos orgánicos complejos desde moléculas inorgánicas simples como el agua y el dióxido de carbono por el proceso de la fotosíntesis. Los hongos, en contraste, todos requieren un suplemento de compuestos orgánicos preformados para su producción de energía y crecimiento. Son descritos como organismos heterotróficos, de aquí que los heterótrofos sean nutridos desde otra parte, es decir, no son capaces de alimentarse por si mismos como hacen las plantas autótrofas. Muchos hongos son encontrados en la oscuridad, hábitat húmedos, pero están universalmente presentes allí donde se encuentra la materia orgánica.
Los hongos pueden ser saprofíticos o parásitos. Los organismos saprofíticos son definidos como aquellos que viven en la materia orgánica en descomposición mediante la degradación de las plantas. Los parásitos obtienen sus nutrientes de animales y plantas vivos, y generalmente causan enfermedades en sus hospedadores. La mayoría de los hongos se encuentra como saprófitos en el suelo, viviendo en material vegetal en descomposición, donde estos juegan un papel vital en el reciclado de la materia orgánica. Los hongos se alimentan por secreción de enzimas hidrolíticas dentro de su medio local.
En cultivos artificiales, muchos hongos pueden crecer en un medio de sales minerales conteniendo una fuente de sales nitrogenadas, que contiene glucosa presente como fuente de carbono pueden manufacturar todas las moléculas orgánicas que requieren para crecer a partir del metabolismo de la glucosa. Otros hongos requieren un suplemento exógeno de vitaminas u otros factores de crecimiento que no pueden fabricar por ellos mismos en orden a crecer en cultivos artificiales. Los factores de crecimiento añadidos son de particular importancia si las células vegetativas en cultivo están por iniciar la esporulación. Los hongos tienen unos requerimientos específicos de elementos traza incluyendo calcio, magnesio, hierro, zinc, cobre y manganeso. No se han encontrado hongos que puedan fijar el nitrógeno atmosférico.
8.9. CLASIFICACIÓN DE LOS HONGOS FITOPATÓGENOS
HONGOS INFERIORES
Clase: MYXOMYCETES (mohos mucilaginosos). Carecen de micelio. Su forma es un plasmodio amorfo y desnudo.
Orden: Physarales. Forman un plasmodio saprofito que produce cuerpos fructíferos costrosos que contienen esporas. Producen zoosporas.
Genero: fuligo, mucílago y physarum producen mohos mucilaginosos en plantas de poca altura. Orden: Plasmodiophorales. Los plasmodios se forman en el interior de las células de las raíces y tallos de las plantas. Producen zoosporas.
Géneros: Plasmodiophora. P. brassicae produce la hernia de las crucíferas. Polymyxa. P. graminis parasita al trigo y a otros cereales. Spongoporas. S. subterránea produce la sarna polvorienta de los tubérculos de papa. Urophlyctis. U. alfalfae produce la verruga de la corona de la alfalfa.
Clase: PHYCOMYCETES (hongos algaceos); hongos inferiores verdaderos.
Subclase: CHYTRIDIOMYCETES.- Tiene un micelio redondeado o alargado que carece de septos.
Orden: Chytridiales. Tienen pared celular pero carecen de un micelio verdadero; la mayoría de ellos forman un rizomicelio y producen zoosporas.
Géneros: Olpidum. O. brassicae parasita las raíces de la col y de otras plantas. Physoderma. P . maydis produce la mancha parda del maíz. Synchytrium. S. endobioticum produce la verruga de la papa. Urophlyctis. U . alfalfae produce la verruga de la corona de la alfalfa.
Subclase: OOMYCETES(mohos acuáticos, royas blancas y mildius). Tienen un micelio alargado. Producen zoosporas en zoosporangios. Las oosporas se forman por la fusión de gametos morfológicamente distintos.
Orden: Saprolegniales. Tienen un micelio bien desarrollado. Las zoosporas se forman en largos zoosporangios cilíndricos que se encuentran fijos al micelio, forman oosporas.
Genero: Aphanomyces ocasión la pudrición de la raíz de varis hortalizas.
Orden: Peronosporales. Los esporangios (por lo común, zoosporangios) se forman en las puntas de las hifas y quedan libres. Forman oosporas.
Familia: Pythiaceae. Géneros: Pythium produce el ahogamiento de las plántulas, pudriciones de semillas y raíces y el tizón algodonoso de los céspedes. Phytophthora. P. infestan produce el tizón tardío de la papa y otras especies producen la mayoría de las pudriciones de la raíz.
Familia: Albuginaceae (royas blancas).
Genero: Albugo. A. candida produce la roya blanca de las crucíferas.
Familia: Peronosporaceae (mildius)
Genero: Plasmopara. P . Vitícola produce el mildiu de vid. Peronospora. P. nicotianae produce el mildiu (moho azul) del tabaco. Bremia. B. lactucae produce el mildiu de la lechuga. Sclerospora. S. graminicola produce el mildiu de las gramíneas. Pseudoperonospora. P. cubensis produce el mildiu de las cucurbitáceas.
Subclase: ZYGOMYCETES (Mohos del pan). Hongos terrestres. Producen esporas asexuales no móviles en esporangios. No forman zoosporas. Su espora de resistencia es una zigospora que se forma por la fusión de un par de gametos morfológicamente idénticos.
Orden: Mucorales producen zigosporas. Las esporas asexuales no moviles se forman en esporangios terminales.
Géneros: Rhizopus produce la pudrición blanda de los frutos y hortalizas. Choanephora. C. cucurbitarum produce la pudrición blanda de la calabaza
HONGOS SUPERIORES
Clase: ASCOMYCETES (hongos de saco). Producen grupos de ocho esporas asexuales, denominadas ascoporas, en el interior del asca.
Subclase: HEMIASCOMYCETES. Con ascas desnudas que no se forman en ascoporas.
Orden: Taphrinales. Las ascas se forman a partir de células ascogenas binucleadas.
Genero: Taphrina. Produce el enrizamiento de las hojas del durazno, abolsamiento del ciruelo, la verruga foliar del roble, etc.
Subclase: EUASCOMYCETES. Las ascas se forman en ascocarpos.
Serie: PYRENOMYCETES (hongos periteciales). Las ascas se forman en cuerpos fructíferos totalmente cerrados (cleistotecios) o en cuerpos fructíferos que presentan una abertura (peritecios).
Orden: Erysiphales (cenicillas). El micelio y los cleistotecios se forman sobre la superficie de la planta hospedera.
Géneros: Erysiphe produce la cenicilla de las gramíneas, cucurbitáceas, etc. Microsphaera; una especie produce la cenicilla de la lila. Podosphaera. P. leucotricha produce la cenicilla del manzano. Sphaerotheca. S. pañosa produce la cenicilla del rosal y del durazno. Unicinula. U. necator produce la cenicilla de la vid.
Orden: Sphaeriales. Los peritecios poseen paredes firmes y colores obscuros.
Géneros: Ceratocystis. C. ulmi produce la enfermedad del olmo holandés. Diaporthe produce el tizón de la vaina del fríjol, la melanosis de los cítricos y la pudrición del fruto de la berenjena. Endothia. E. parasitica produce el tizón del castaño. Glomerella. G. cingulata produce muchas antracnosis y la pudrición amarga del manzano. Gnomonia produce la antracnosis o mancha foliar de varias plantas. Rosellinia produce las enfermedades de la raíz de la vid y de los árboles frutales. Valsa produce el cáncer del durazno y de otros árboles. Xylaria produce el cáncer de los árboles y la pudrición de la madera.
Orden: Hypocreales. Los peritecios son de colores claros, rojos o azules.
Géneros: Claviceps. C. purpúrea produce el cornezuelo del centeno. Gibberella ocasiona la pudrición del pie o tallo del maíz y de pequeños granos. Nectria produce el cáncer del tallo y ramas de los árboles.
Serie: PSEUDOSPHAEROMYCETES (hongos ascostromaticos). Presentan estromas en forma de peritecio que presentan ascas en cavidades separadas o en grandes cavidades.
Orden: Myriangiales. Con cavidades dispuestas a varios niveles y que contienen ascas individuales. 7 Genero: Elsinoe produce las antracnosis de la vid y de la frambuesa y la saran de los cítricos.
Orden: Dothideales. Con cavidades dispuestas en una capa basal, las cuales contienen muchas ascas. Los peritecios carecen de pseudoparafisas.
Géneros: Dibotryon. D. morbosum produce la nodulacion negra de las ramas en cerezos y ciruelos. Dothidella. D. ulei ocasiona la mancha foliar de los árboles del caucho. Gugnardia. G. bidwelli produce la pudrición negra de las uvas. Mycosphaerella produce las manchas foliares de muchas plantas.
Orden: pleosporales. Con cavidades dispuestas en una capa basal, las cuales contienen muchas ascas. Los peritecios presentan pseudoparafisas.
Géneros: ophiobolus. O. graminis produce la enfermedad del pie de trigo. Physalospora. P. obtusa produce la pudrición negra de la manzana. Venturia. V. inaequalis ocasiona la roña de la manzana
Serie: DISCOMYCETES (hongos de copa). Las ascas se forman en la superficie de apotecios carnosas en forma de copa o de plato.
Orden: Helotiales. Las ascas liberan sus esporas a través de una perforación apical o circular.
Géneros: Coccomyces. C. hiemalis produce la mancha foliar del cerezo. Diplocarpon. D. rosae produce la mancha negra de las rosas. Lophdermium produce el tizón de las agujas del pino. Monilinia. M. frucyicola produce la mancha parda de los frutos de hueso. Rhytisma. R. acerium produce la mancha alquitranada de las hojas del arce. Sclerotinia. S. sclerotiorum produce la pudrición blanda aguanosa de las hortalizas.
Orden: pezizales. Las ascosporas se liberan a través de una estructura en forma de tapa o capsula que se localiza en la punta del asca.
Genero: pseudopeziza. P. medicaginis produce la mancha foliar de la alfalfa.
Clase: FUNGI IMPERPECTI O DEUTEROMYCETES (hongos asexuales). Carecen de estructuras o reproducción sexuales o no se sabe que las presenten.
Orden: Sphaeropsidales. Las esporas asexuales se forman en picnidios.
Géneros: Ascochyta. A. pisi produce el tizón del tallo de la frambuesa. Cytospora ocasiona el cáncer del durazno y otros árboles. (Valsa representa su etapa sexual). Diplodia. D. zeae produce la pudrición del talo y la mazorca del maíz. Phoma. P. lingam ocasiona la pierna negra de las crucíferas. Phomopsis produce al tizón y el cáncer del tallo de varios árboles. Phyllosticta produce las manchas foliares de muchas plantas. Septoria. S. apti produce el tizón tardío del apio.
Orden: Melanconiales. Las esporas asexuales se forman en un acervulo.
Géneros: Colletotrichum ocasiona la antracnosis de muchas plantas de cultivo. 8 Coryneum: C. beijerincki produce el tizón de los frutos de hueso. Cylindrosporium produce manchas foliares en muchas clases de plantas. Gloeosporium muy parecido (si no idéntico) a Colletotrichum; produce antracnocsis en muchas plantas. Marssonina ocasiona el tizón de las y hojas del álamo, la quemadura de las hojas de fresa y la antracnosis de los nogales. Melanconium. M. fuligenum produce la pudrición amarga de la vid. Sphaceloma produce la antracnosis de la vid y de la frambuesa y la sarna de los cítricos del aguacate. Orden: Moniliales. Las esporas asexuales se forman sobre las hifas (o en su interior) del hongo que se encuentran expuestas libremente a la atmósfera.
Géneros: Alternaría produce manchas foliares y tizones en muchas plantas. Aspergillus produce la pudrición de las semillas almacenadas. Botrytis. B. cinerea produce el moho gris y los tizones de muchas plantas. Cercospora; una especie de este genero produce el tizón temprano del apio. Cladosporium. C. fulvum produce el moho de las hojas del tomate. Fusarium produce el marchitamiento y la pudrición de la raíz de muchas plantas anuales, así como el cáncer de árboles forestales. Fusicladium produce la roña de la manzana (venturia representa su etapa sexual). Graphium. G. ulmi produce l enfermedad del olmo holandés (Ceratocystis representa su etapa sexual). Helminthosporium produce el tizón de los cereales y enfermedades de los céspedes. Penicillium produce la pudrición de los frutos y otros órganos carnosos debido a los mohos azules. Phymatotrichum. P. omnivorum produce la pudrición de la raíz del algodonero y otras plantas. Pyricularia produce el tizón del arroz y la mancha gris foliar de los céspedes. Strumella produce el cáncer del roble. Thielaviopsis. T. basicola produce la pudrición negra de la raíz del tabaco. Verticillium produce la marchitez de muchas plantas anuales y perennes.
Orden: Mycelia Sterilla. No se ha observado o es muy poco frecuente la formación de esporas asexuales o sexuales en este grupo de hongos. Géneros: Rhizoctonia produce las pudriciones de la raíz de la corona de las plantas anuales y mancha parda de los céspedes (su etapa perfecta corresponde a Thanatephorus).
9. IDENTIFICACIÓN, ESTRUCTURAS VEGETATIVAS Y REPRODUCTIVAS DE LOS HONGOS FITOPATÓGENOS.
9.1. TÉCNICAS PARA LA IDENTIFICACIÓN DE LOS HONGOS
Para la identificación de hongos fitopatógenos es necesario la observación de sus estructuras somáticas y reproductivas. Mediante la técnica de cámara húmeda y/o aislamiento es posible inducir la aparción de estas estructuras. La observación de las características de las estructuras producidas y el uso de claves taxonómicas son necesarias para determinar el género y la especie del hongo patógeno.
9.2. ESTRUCTURAS VEGETATIVAS
Plasmodio: Se refiere al cuerpo o soma vegetativo de algunos hongos inferiores, el cual está constituído por una masa multinucleada, sin pared celular. Son escasos los hongos fitopatógenos que poseen soma vegetativo de tipo plasmodial. Micelio: La mayoría de los hongos poseen cuerpo filamentoso provisto de pared celular. . A los filamentos que constituyen el cuerpo o soma vegetativo se les denomina hifas. Al conjunto de hifas se le denomina micelio. Cuando las hifas no presentan septas, el micelio es denominado cenocítico o no tabicado y cuando las presenta el micelio se dice que es tabicado.
Esclerociosos: Los esclerotos son estructuras de resistencia formados por compactación de hifas. Estos esclerociosos pueden tener diferentes formas y tamaños. A continuación se muestran diferentes tipos de esclerociosos:
9.3. ESTRUCTURAS REPRODUCTIVAS
HONGOS INFERIORES Y PSEUDOHONGOS
Cuerpos fructíferos representativos de los hongos inferiores y pseudohongos: Los hongos inferiores se caracterizan por poseer micelio cenocítico. Según su reproducción sexual estos hongos pueden pertenecer a dos diferentes clases:
Clase: Oomycetes: (Ficomicetes) Esta clase de hongos sexualmente producen oosporas, las que se originan por la unión de dos gametos diferentes, el oogonio y el anteridio. Estas oosporas son esféricas y de pared gruesa. Estos hongos se reproducen asexualmente producen esporas flageladas, denominadas zoosporas. Estas zoosporas se encuentran contenidas en cuerpos fructíferos denominados zoosporangios.
Clase: Zigomycetes: Esta clase de hongos Produce esporas asexuales no móviles contenidas en cuerpos fructíferos denominados esporangios. Sexualmente producen esporas denominadas zigosporas
10. ESTRUCTURAS REPRESENTATIVAS DE LOS HONGOS SUPERIORES.
10.1. ESTRUCTURAS REPRESENTATIVAS DE LA CLASE ASCOMYCETES.
La clase ascomycetes se caracteriza por poseer micelio tabicado y producir esporas de origen sexual denominadas Ascosporas. Estas ascosporas se producen dentro de sacos llamados ascas. Las ascas pueden encontrarse en forma libre o contenidas en cuerpos fructíferos. Los cuerpos fructíferos pueden ser de dos tipos: Apotecios y Cleistotecios. A continuación se muestran ejemplos de estas estructuras:
Ascas libres:
Ascas en Apotecios:
Los apotecios son cuerpos fructíferos con forma de copa. Normalmente de colores marrones claros, se pueden producir a partir de la germinación de esclerotos o del mismo material vegetal invadido por el hongo. Algunos alcanzan más de un centímetro de diámetro. Un corte de la copa permite observar bajo el microscopio la presencia de ascas conteniendo ascosporas.
Acas en Peritecio:
Los peritecios son cuerpos fructíferos con forma de pera y con una abertura para la salida de las ascosporas. Generalmente se producen sobre el tejido vegetal colonizado por el hongo. A simple vista pueden observarse como puntuaciones negras.
Ascas en Cleistotecio:
Los Cleistotecios son cuerpos fructíferos con forma con forma esférica y que no poseen abertura para la salida de las ascosporas, por lo tanto deben romperse para que estas se liberen. Generalmente se producen sobre el tejido vegetal colonizado por el hongo. A simple vista pueden observarse como puntuaciones negras.
11. ESTRUCTURAS REPRESENTATIVAS DE LOS HONGOS SUPERIORES
11.1. CLASE BASIDIOMYCETES.
La clase basidiomycetes se caracteriza por por tener micelio tabicado y reproducirse sexualmente mediante la producción de basidioporas. Estas son producidas exógenamente sobre una estructura llamada basidio. Los basidios pueden ser septados o no.
El micelio primario, monocariótico. Esta fase suele ser corta, ya que pronto ocurre la somatogamia o fusión de hifas y se obtiene un micelio secundario, dicariótico, que crece mediante fíbulas . Este micelio es el más abundante en la naturaleza. En algunos casos de hongos micorrizógenos, puede ocupar varias hectáreas, pesar bastantes toneladas y tener una edad de varios milenios. Los septos son complejos , ya que presentan un poro central (dolíporo) rodeado de una serie de membranas (parentosoma). En ocasiones, el micelio puede formar cordones o rizomorfos . También se dan casos en que el micelio es un diploide estable, como en Armillaria.
Aunque no es tan corriente como la somatogamia, el micelio secundario puede originarse por un proceso de espermatización. Algunos hongos producen oídios (es importante no confundirlos con las conocidas cenizas, que ya se vieron al tratar los ascomicetos) en unas hifas especiales, los oidióforos. Estos oídios están rodeados de una gotita de mucus, para su dispersión por el agua o los insectos. Tarde o temprano, contactarán con una hifa compatible y ocurrirá la dicariotización.
En cuanto a la compatibilidad sexual, las especies homotálicas son raras, y las heterotálicas abundan. De estas últimas, casi el 25% regulan el tipo de sexo mediante alelos en un único locus (heterotalismo unifactorial o bipolar). Es el caso de royas, casi todos los carbones y algunos yesqueros y levaduras. En cambio, la mayoría de basidiomicetos regulan el tipo sexual mediante más de un par de genes, localizados en diferentes cromosomas (heterotalismo bifactorial o tetrapolar, aunque, a pesar de lo que indica el nombre, puede implicar a más de dos pares de genes).
El micelio secundario puede reproducirse asexualmente por medio de conidios (raro), por gemación (en levaduras y carbones), por esporas especiales (en royas), e incluso mediante esclerocios. Como en otros hongos, la fragmentación del micelio es un método de dispersión frecuente.
Sin embargo, lo más típico es la reproducción sexual (aunque existen especies que no la presentan). El micelio secundario puede agruparse en «tejidos» especializados plectenquimáticos, aún dicarióticos, y el micelio se denomina entonces terciario. Da lugar a cuerpos fructíferos, los basidiocarpos o basidiomas, algunos de los cuales son bastante grandes, hermosos y comestibles: las conocidas setas. En el campo es frecuente encontrar grupos de setas dispuestas en anillo, los corros de brujas , debido al crecimiento en todas direcciones del micelio y la aparición de basidiocarpos en la periferia. En cambio, otros basidiocarpos son muy diferentes a las setas : leñosos, gelatinosos, inconspicuos
Los basidios se disponen sobre o dentro del basidiocarpo, a menudo en una capa fértil, el himenio . Entre los basidios pueden existir estructuras estériles, como los basidiolos(recuerdan a los basidios, pero que no producen esporas) o los cistidios, de mayor tamaño y muy útiles como carácter taxonómico. Cada basidio dará lugar a 4 (pero pueden ser 2, 8 ó más) basidiósporas externas.
11.2. CARACTERÍSTICAS
Este filo es el más evolucionado y el más conocido pues comprende numerosos y variados tipos de hongos. Cuando son de carácter heterotálico, el micelio primario sufre dicariotizacion (somatogamia o espermatización) produciendo hifas dicarióticas que corresponden al micelio secundario. En los hongos de carácter homotálico una basidiospora produce el micelio dicariótico. Hay presencia de quitina en las paredes celulares, y aparecen unas estructuras llamadas fíbulas, muy parecidas a los uncínulos de los ascomicetos.
11.3. TIPOS
Heterobasidiomycetes. Cuando los basidios están septados y muy divididos. Las esporas son resistentes por poseer una pared muy gruesa. Presentan más de un tipo de conidio.
Homobasidiomycetes. Cuando los basidios son más uniformes y no septados con forma claviforme. En estos la basidiospora germina formando la hifa. Los órdenes son tremelales, uredinales y ustilaginales.
12. ESTRUCTURAS REPRESENTATIVAS DE LOS HONGOS SUPERIORES
12.1. CLASE DEUTEROMYCETES
Esta Clase incluye a los hongos superiores (micelio tabicado) a los que no se les conoce la reproducción sexual. En algunos casos por no tener reproducción sexual, o si la tienen esta se produce rara vez, o simplemente porque no se le conoce aún. Estos hongos se reproducen de forma asexual formando esporas denominadas 18 conidios. Estos conidios pueden producirse en forma libre o dentro de cuerpos fructíferos. Los conidios pueden tener diferentes formas y tamaños. Pueden ser hialinos u obscuros. Pueden ser unicelulares o multicelulares. Pueden estar sueltos o agrupadosnen ramilletes o cadenas.
Conidios en cuerpos fructíferos:
Acérvulos: Los acérvulos son estructuras abiertas sobre la superficie del vegatal que dejan expuestos a los conidios.
Picnidios:
Los picnidios son curpos fructíferos que poseen una abertura por la que liberan los conidios producidos en su interior.








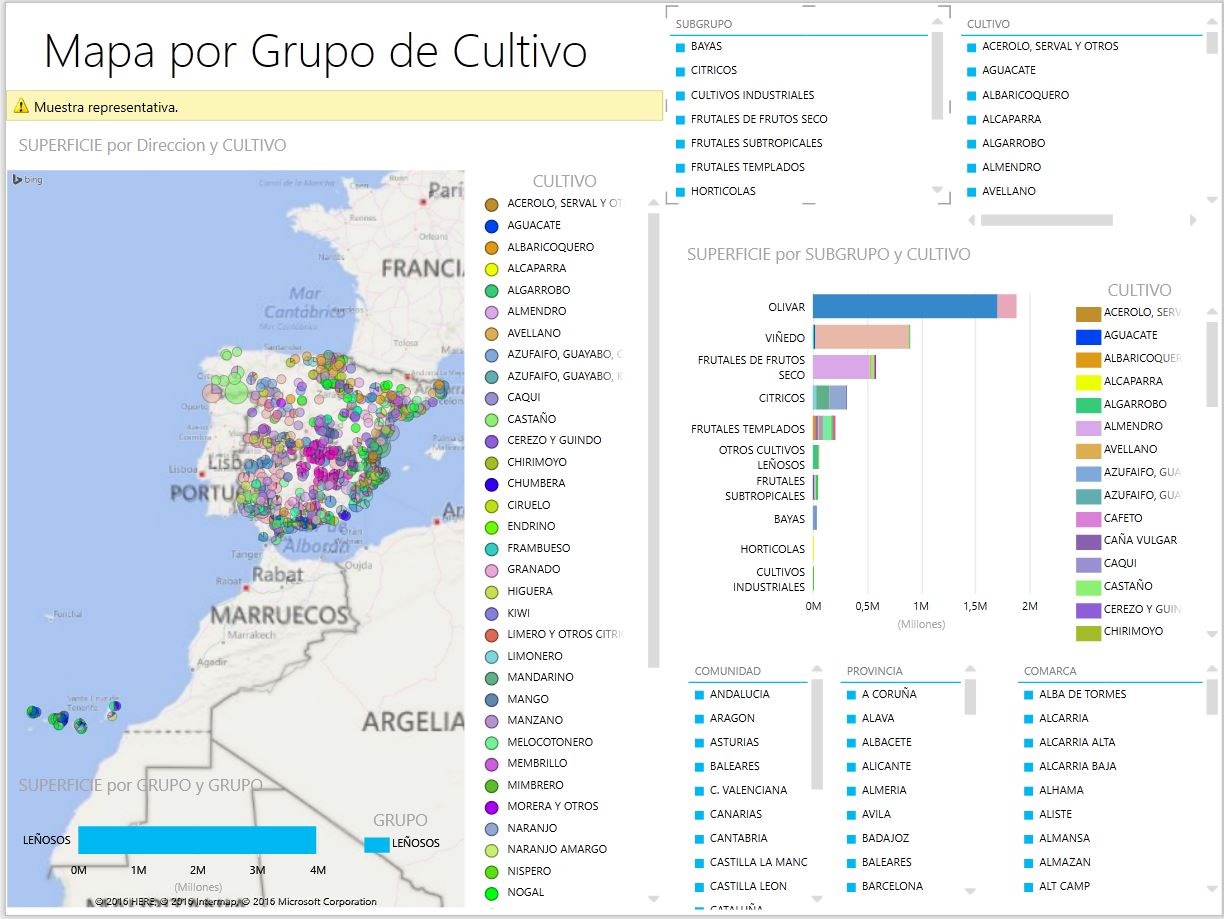








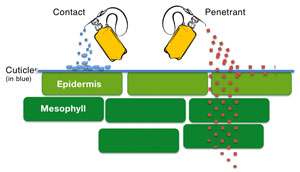
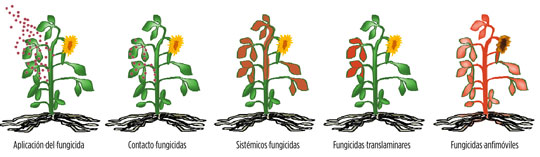
 Edición
Edición



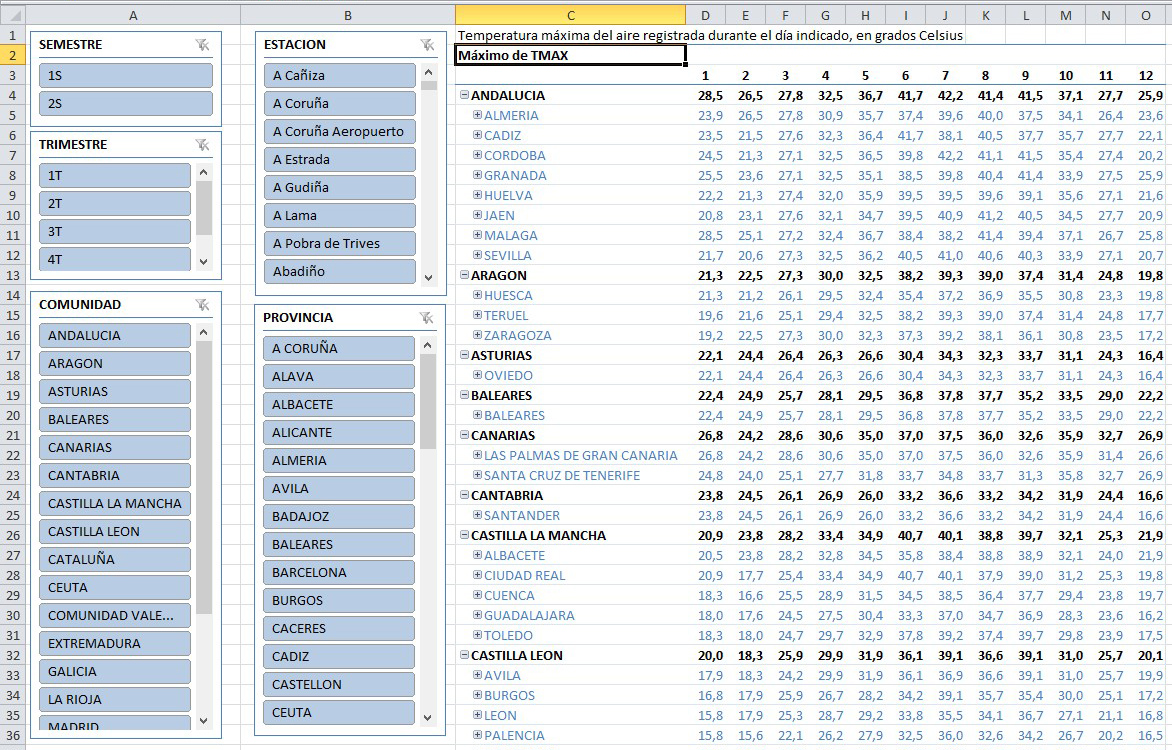




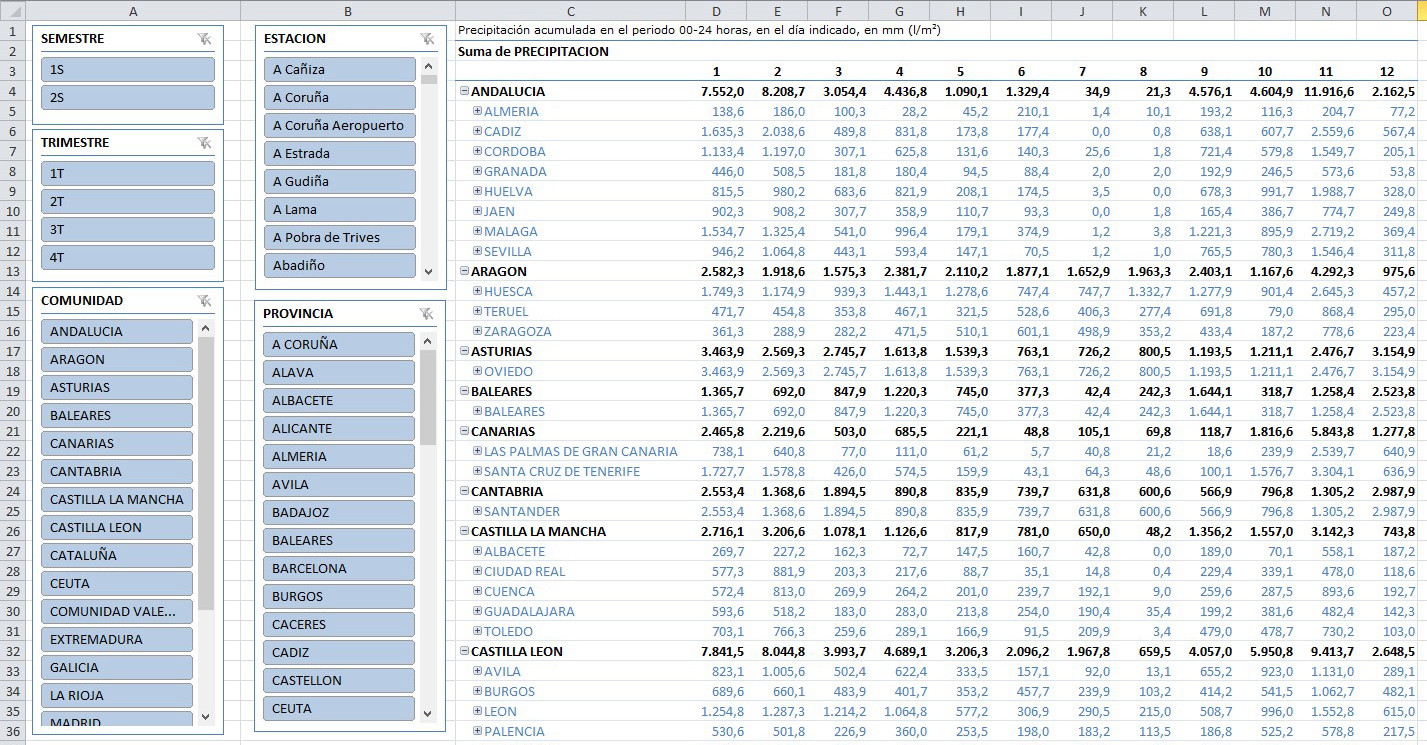


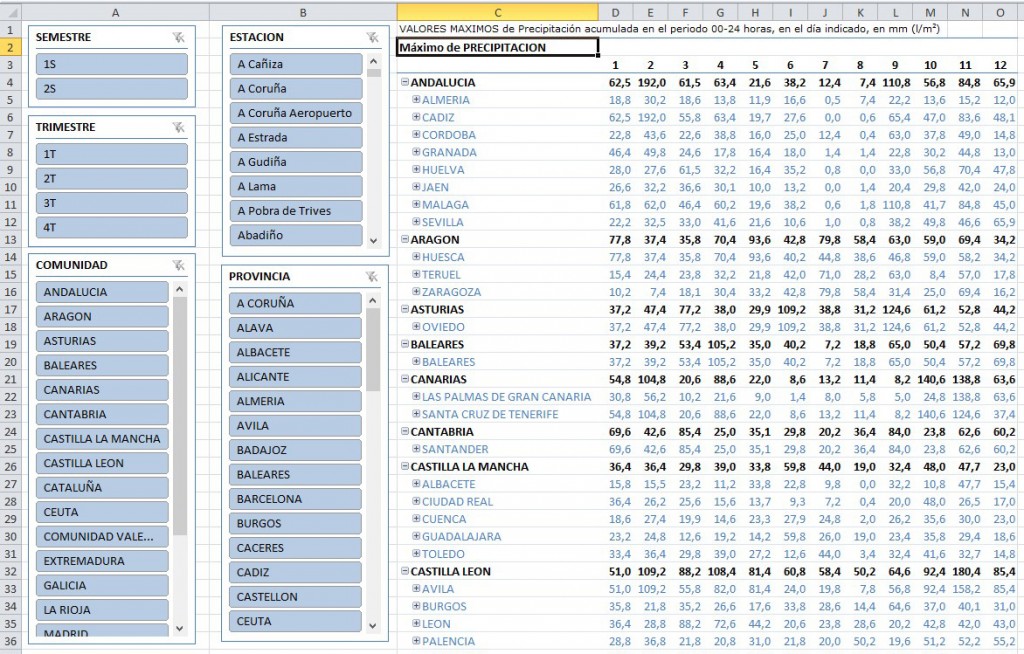
{kind=link}